
kukkivat


1. kukkapohja (Receptakulum)
2. verholehdet (verholehtiset)
3 terälehteä (terälehtiä)
4. heteet (heteet)
5. leima (survin)
Kukinta kasvi on haarautumaton, lyhyt verso rajoitettu kasvu, joka voidaan havaita sen jälkeen, kun kukinnan juoksuaika , lehdet, jotka ovat suoraan tai epäsuorasti, että palvelu on sukupuolisen lisääntymisen : välillisesti suoja- tai houkutella elinten ( kukka kirjekuori ), suoraan lisääntymiselinten ( heteet ja karvat ) muodostuminen.
Usein vain angiospermien kukka ymmärretään kukan alla , koska ne ovat kapeammassa mielessä kukkivia kasveja. Siitä huolimatta voimisteluissa on myös kukkia tai kukintoja .
Angiosperm -kukka koostuu seuraavista osista (mutta kaikkia ei tarvitse esittää): kukka -akseli (kukkapohja), josta kukka -elimet syntyvät: kukka -kirjekuori (perianth) perigonina tai erotettuna verhiöksi ja terälehtien kirjekuori , hedeistä valmistettu androeceum ja karpeleista valmistettu gynoeceum . Hedelmät muodostavat siitepölyn, joka pölytyksen aikana saavuttaa kappelin leimautumisen, itää siellä ja sijaitsee hedelmöitetyissä kappelin munasoluissa , tarkalleen munasolussa ( hedelmöitys ). Hedelmöitetystä munasolusta kehittyy kasvien alkio , munasolusta siemen ja kaikista kukan muista osista tulee hedelmiä .

Kukkia, jotka sisältävät vain karpeleita, kutsutaan puhekielessä "naisiksi", niitä, jotka sisältävät vain heteitä, kutsutaan "uroksiksi" ja kukkia, joissa on sekä heteitä että karvoja, kutsutaan biseksuaaleiksi tai hermafrodiittikukiksi (ks. Alla: Seksuaalisuus ).
Määritelmät
Kukan laaja määritelmä (keski -yläsaksasta / vanhasta yläsaksalaisesta bluotista, jossa on monikko blüete ), kuten se usein esiintyy kasvitieteellisessä erikoiskirjallisuudessa, sisältää osittain nimenomaisesti tai epäsuorasti alastomien kukkia . Vain harvoin se on implisiittisesti rajoitettu selkärankaisiin nimeämällä määritelmän osat. Vuonna Strasburger oppikirja kasvitieteen , kaikki siemenkasvit kutsutaan niiden kukkia, vaikkakin rajoituksia: ”in spermatophytina The sporophylls ovat lähes aina lyhyen ituja rajalliseen kasvuun. Tällaisia [...] rakenteita voidaan kutsua kukiksi, vaikka tätä termiä käytetään enimmäkseen vain pääasiassa hermafrodiittisorofyllitasoihin, joita ovat siemenet ja mahdollisesti läheisesti liittyvät fossiiliset siemenkasvit. ” Biologian sanastossa määritellään kukka lyhyesti myös” lyhyt itu, jonka kasvu on vähäistä ja joka kantaa sporofylliä ".

Esimerkki naaras- ja uroskukinnosta sykistä

Esimerkki voikukan siittiöiden naaraspuolisesta kukinnosta (tässä mänty )

Harvinaisen männyn naaraspuolinen kukinto suurentunut

Esimerkki urospuolisesta kukinnasta voimistelussa (tässä harvinainen mänty)

Esimerkki kukka eudicotyledons kanssa verholehdet , terälehdet , emi ja heteet (tässä niitty kurjenpolvi )

Esimerkki kukinnan yksisirkkaisten kanssa Perigon , leima ja heteet (tässä Palo Lilja )

Esimerkki kukinnosta makeissa ruohoissa






.jpg)
Fossiilisia edustajia kukkien muodostavien siemenkasvien ulkopuolella ovat Bennettitales ja Caytoniales . Kukan määritelmä vastaa myös karhun sammalta ja korttia sporofyllitasoja , mutta niitä ei käsitellä tässä tarkemmin.
Englanninkielisissä maissa kukka (kasvitieteellisesti: kukka ) määritellään yleensä suppeammin angiospermin kukana, esim. Esimerkki: "lyhennetty, hermafrodiittinen lisääntymiskannan akseli, jonka kasvu on rajoitettua ja joka koostuu megasporangiasta (karselit) ja mikrosporangiasta (heteet) ja steriilistä kukan kirjekuoresta, jossa on vähintään yksi steriili, litteä elin."
morfologia
Angiospermissä yksittäiset kukkaelimet on järjestetty kukkaan kiinteässä järjestyksessä alhaalta ylöspäin tai, koska kukka -akseli on puristettu, ulkopuolelta sisäänpäin. Tässä järjestyksessä niitä käytetään myös ontogeneettisesti (kukkien muodostumisen aikana).
Kukka -akseli
Terälehdet kiinnittyvät kukka -akseliin (kukkapohjaan tai astiaan). Se voidaan puristaa, venyttää ja muotoilla eri tavalla. Se voi kerätä yksittäisiä tai useita kierteitä interkalaaristen meristeemien avulla : Esimerkiksi kun Kapernstrauch on gynoecium nostettu ( Gynophor ) intohimo kukka on gynoecium ja Androeceum nostettu ( Androgynophor ), ja monissa neilikan kasveissa ovat kruunu, Androeceum gynoecium nostettu ( Anthophor ).
Keskuksen ympärillä olevia rengasmaisia alueita voidaan myös nostaa. Yksi esimerkki on stamen-Corollentubus niin monta komposiitti kukkia on tyypillistä, tai kuppi-Kronbecher monia cucurbits . Levennetyn kukka akseli kutsutaan kukan kehtoa (hypanthium).
Jos kaikki osat paitsi munasarja nostetaan ylös, syntyy kukkaputki, joka kantaa kukkakuoren ja heteet. Tuloksena on perigynous kukkakuppi. Munasarjaa kutsutaan sitten keskellä olevaksi. Ylempi munasarja seisoo vapaasti kukka -akselilla. Jos karppien ulkopuolet sisältyvät kukkaruukkuun, syntyy alisteinen munasarja, mikä tarkoittaa, että se on kukkakuoren alapuolella, esimerkiksi ruusujen tapauksessa .
Perianth
Perianth (perianth) ovat steriilejä levyjä , mutta kuuluvat selkeästi kukkia. Kukan kirjekuoren muodostavat vain Gnetopsida ja angiospermit . Jos kaikki kukinta verhouksen levyt tasaisesti suunniteltu (Homoiochlamydeisch), kukinta kirjekuori kutsutaan perigon tai yksinkertainen perianth , lehdet kuten tepals (esimerkiksi kanssa tulppaani ). Perigoni voi koostua yhdestä ympyrästä (haplo- tai monoklamyylinen), kahdesta tai useammasta ympyrästä tai useista ruuvikierroksista; useita perigoneja . Kaksinkertainen perianth (di- heterochlamydeic kukat) koostuu erilaisten suojuslehtien. Ulkoiset, enimmäkseen vihreät, ovat sepals (sepals) ja muodostavat verhiön (verhiö), sisäiset, usein silmiinpistävän väriset lehdet ovat terälehtiä ja muodostavat seppeleen. Kukan kirjekuoren evoluution alkuperä tulkitaan kannateista tai hedeistä sukulaisryhmästä riippuen .
Mikrosporofyllit


Mikrosporofyllit ovat lehtiä, jotka kantavat sporangiaa. Mies mikrosporeja on muodostettu , että sporangioita (siitepöly pusseja) . Mikrosporofyllit kehittyvät hyvin eri tavalla siemenkasviryhmästä riippuen, eikä homologiaa lehtien kanssa (φύλλον phýllon "leaf") ole täysin selvitetty. Siksi niitä voitaisiin kutsua myös neutraalimpiksi mikrosporangian kantajiksi. In neidonhiuspuu , microsporophyll on varsi, jossa on kaksi siitepöly pusseja roikkuu kärjestä; Liian monet mikrosporofyllit ovat ruuvimaisia kukka-akselilla. Että havupuiden , mies kukka on kartion kaltainen ja koostuu useista microsporophylls, yleensä järjestetty kierre; jokaisessa mikrosporofyllissä on 2–20 siitepölypussia. Vuonna käpypalmuja (käpypalmuja), jotka ovat limittäisen ja microsporophylls viidellä 1000 siitepöly pusseja; mikrosporofyllit ovat uroskukan ruuvin muotoisia. Vuonna Welwitschia , kuusi microsporophylls jotka ovat kasvaneet yhdessä tehdä kolme siitepöly pusseja kullekin. In Gnetum vain microsporophyll on yksi tai kaksi terminaalista siitepöly pusseja. Vuonna ephedra The microsporophyll usein haarautuu ja kannattaa kahta kohteeseen kahdeksaan useimmiten kahden intergrown siitepöly pusseja.
Angiospermien mikrosporofylli kutsutaan hedeiksi. Hede on yleensä filamentissa (filamentti) ja pölypussi jaettuna (anher). Jälkimmäinen koostuu kahdesta laskurista, joissa molemmissa on kaksi siitepölysäkkiä ja jotka on liitetty liittimellä. Heteiden kokonaisuutta kutsutaan Androeceumiksi . Heteitä kukkaa kohti on yksi ja noin 2000 kappaletta.
Megasporofyllit



Megasporofyllit sisältävät siemenkasveille ominaisia munasoluja , joiden sisällä ovat varsinaiset naisen lisääntymiselimet, jotka sisältävät kasvien munasoluja. Megasporofylejä voitaisiin kuvata myös neutraalimmin megasporangian kantajina. Vain angiospermissä megasporofylejä kutsutaan karpeleiksi .
Sykadien munasolut ovat selvästi lehdissä, ne ovat phyllospor. Kattilakiveä, kuten tai kilpi-muotoinen megasporophyll on selvästi stalked ja on kaksi ovuleina alareunaan ja terän , jossa on Cycas on jopa kahdeksan ovuleina pitkin rachis . Havupuilla on 1-20 munasolua tasaisen siemenvaa'an pinnalla. Siemenvaaka on muokattu lyhyt ampuma. Havupuiden munasolut ovat harvoin lyhyiden versojen lopussa, kuten marjakuusi ( Taxus ). Ginkgo muodostaa kaksi munasolua haarautuneen varren kärkeen. Gnetopsida muodostavat vain yksi terminaali munasolun kohti kukka.
Angiospermsissä munasolut ovat Carpel ( carpel ), joten nimi "angiosperms". Kaarleiden kokonaisuus muodostaa gynoeceumin . Karpelien lukumäärä kukkaa kohden on 1–2000. Karppeli on yleensä jaettu varsivyöhykkeeseen ja putkimaiseen vyöhykkeeseen, onttoon alueeseen, jossa munasarjat sijaitsevat (kutsutaan myös munasarjaksi). Munasolut kiinnittyvät istukkaan . Tämän yläpuolella on usein kynä , jolla arpi sijaitsee, pinta siitepölyn saamiseksi. Arpi on liitetty munasoluihin siirtokanavalla tai kudoksella. Karvat voivat olla vapaita (apokarpet tai chorikarpes gynoeceum) tai fuusioituneet yhteen (koenokarpit). Kasvaneen gynoeceumin tapauksessa puhutaan leimasta (survin). Steriileissä kukissa on korkeintaan vähentynyt hede ja kaari, ja ne toimivat usein näyttölaitteena .
Nektarit
Nektarit ovat rauhasia, jotka tuottavat nektaria vieheenä pölyttäjille. Tämä koostuu yleensä sokeriliuoksesta , joka voi sisältää myös proteiineja , aminohappoja ja vitamiineja . Öljyjä muodostuu harvemmin (esimerkiksi löysässä ajassa , Lysimachiassa ). Nektaria voidaan muodostaa kukka -kirjekuoresta, hedeistä, munasarjasta, kukka -akselista, mutta myös varsinaisen kukan ulkopuolelta (extrafloral).
Sijainti ja järjestely
Terälehtiä voi vaihdella sen mukaan, arkki asentoon ( lehtiasento ) voi olla sijoitettu whorled (syklisesti) akselilla ydin schraubig (asyklinen) tai. Kierteistä järjestelyä pidetään laajalti primitiivisenä piirteenä, ja siihen liittyy usein suurempi terälehtien määrä. Kahden peräkkäisen lehden välinen kulma on yleensä lähellä kultaisen osan kulmaa , noin 137,5 °.
Usein vain osa terälehdistä on ruuhkaisia, kun taas sisemmät terälehdet on järjestetty pyörteeksi. Nämä kukat tunnetaan spirosyklisinä tai hemisyklisinä . Siirtyminen ruuhkaisesta pyörteeseen johtuu siitä, että kuten Yulan -magnolian ( Magnolia denudata ) kohdalla, lehtien muodostumisessa on tauko jokaisen kolmen lehden jälkeen, jolloin muodostuu noin kolminkertaisia pyörteitä. Lehtien muodostumisen kierrejärjestys näkyy myös monien pyörteisten kukkien kehityksessä, mutta ympyrän lehdet ovat niin lähellä toisiaan, että muodostuu pyörteitä.
Pyöreässä asennossa on vain yksi terälehti jokaisessa pyörteessä tai ympyrässä. Pyörivä asento kulkee käsi kädessä myös terälehtien (oligomeeristen ympyröiden) määrän vähenemisen kanssa. Useimmissa pyörtetyissä hermafrodiittikukissa on neljä (tetrasyklisiä) tai viisi (pentacyclic) pyörteitä: sepal, terälehti, yksi tai kaksi heteitä ja kappelin pyörteitä. Useimmissa angiospermeissä lehtien määrä pyöriä on myös kiinteä; vastaavasti puhutaan kahden, kolmen, neljän tai viisinkertaisista kukista. Jos kaikilla ympyröillä on sama määrä linkkejä, kukka on isomeerinen (kuten tulppaanin tapauksessa), jos näin ei ole, kukka on heteromeerinen.
Nikamien lehdet, jotka seisovat päällekkäin, ovat yleensä raolla (vuorotteluperiaate), eli seuraavan sisäpyörän raajat ovat edellisen pyörteen kahden jäsenen välisessä raossa. Pentasyklisten kukkien kanssa tapahtuu usein, että hedeen sisäpiiri työntyy ulospäin munasarjasta ja siten tulee ilmeisesti ulommaiseksi. Tätä ilmiötä kutsutaan obdiplostemonyksi . Kaikki terälehdet voivat olla enemmän tai vähemmän fuusioituneita oman lajinsa tai muiden kanssa.
Jos kukka voi asettaa enemmän kuin kolme symmetriatasoa, se on säteittäinen symmetria (kuten tulppaani ). Kahden symmetriatason tapauksessa se on epäsymmetrinen , esimerkiksi Dicentran tapauksessa . Zygomorfisilla (dorsiventral) kukilla on symmetriataso . Yleensä tämä symmetriataso sijaitsee kukan keskitasossa (kulkee kannen ja varren akselin läpi ), kuten minttuperheessä , harvemmin kohtisuorassa keskitasoon, esimerkiksi leivän kannassa . Epäsymmetrisillä kukilla ei ole symmetriatasoa. Tarkkaan ottaen tämä sisältää myös ruuvinmuotoiset kukat, joita kuitenkin yleensä pidetään säteittäisenä symmetriana.
Kukkaosien suhteet toisiinsa voidaan esittää kukkakaavioissa graafisesti tai kukkien kaavoina .
Usein useita tai monia yksittäisiä kukkia yhdistetään kukintojen muodostamiseksi .
Seksuaalisuus
Angiospermien alkuperäinen muoto ovat hermafrodiittikukat . On myös yksisukukukkia ("erilliset sukupuolet"), vastakkaisen sukupuolen elimet ovat yleensä läsnä myös alkeellisina: On staminaattisia ("uros") ja pistillate ("naaras") kukkia. Unisexual kukilla varustetut kasvit ja klaanit voivat olla yksitoikkoisia ( yksikasvit , uros- ja naaraskukkia yhdellä kasvilla) tai kaksikielisiä ( kaksikieliset , uros- ja naaraskukat eri kasveilla). Moniavioisissa kasveissa on sekä hermafrodiitti- että unisexual -kukkia.
Kukkien induktio
Kukkien induktio on kasvin tai meristeemin kääntyminen vegetatiivisesta kasvusta kukkien muodostumiseen. Liipaisimet voivat olla sisäisiä (endogeenisiä) tai ulkoisia (eksogeenisiä) tekijöitä. Induktio on peruuttamaton, joten sitä ei voi peruuttaa tai pysäyttää. Merkittäviä signalointireittejä on neljä:
- Endogeenisiä tai autonomisia laukaisijoita on läsnä, mutta niiden luonnetta ei tunneta. Monet lajit tarvitsevat kuitenkin tietyn iän tai koon ennen kukintaa, kuten monet puut. Tapauksessa, että herne , kukinnan aika määräytyy geneettisesti lajikkeesta riippuen. Myöhään kukkivat ekotyyppien lituruohon ( Arabidopsis thaliana ) muodossa kukkia itsenäisesti tietyn iän, ellei ollut induktion ympäristötekijät etukäteen. Siirtyminen kukkien muodostumiseen tapahtuu aktivoimalla heterokronisia geenejä, kuten FCA (Flowering Control Arabidopsis).
- Useimpien kasvien kukinnan ajoitus määräytyy ulkoisten tekijöiden mukaan. Tärkeimmät tekijät ovat päivän pituus ja kylmät jaksot ( vernalisointi ). Päivän pituuden suhteen erotetaan lyhyen päivän ja pitkän päivän kasvit, jotka vaativat tiettyjä päivän pituuksia kukinnan aikaansaamiseksi. Fytokromi järjestelmä lehtien näyttelee roolia reseptorin , samoin cryptochrome . FKF1 -proteiini toimii myös fotoretseptorina ja mittaa päivän pituuden. Nämä järjestelmät vaikuttavat Constans -geeniin (CO), jonka ilmentyminen yksinään riittää käynnistämään kukkien induktion. Kukka hormoni Florigen oletettiin pitkään kuin signaalin lehdet kukkia, mutta sen identiteetti jäänyt tuntemattomaksi pitkään. FT -geeni (katso alla) on ehdokas tähän, koska vaikka se transkriboidaan lehtiin, se toimii kukkien meristeemeissä.
- Monocarpic kasveja erityisesti (joka vain kukkivat kerran) vaativat idunviritys pidempi kylmä aikana, voidakseen kukkia. Tunnettuja geenejä, jotka aktivoituvat kylmänä, ovat Frigida (FRI) Arabidopsiksessa ja VRN1 ja VRN2 talvivehnässä. Niiden geenituotteet estävät FLC -geeniä, mikä puolestaan estää kukkien muodostumista. Yhteenvetona voidaan todeta, että FRI tai VRN mahdollistavat kukkien muodostumisen, vaikka niitä ei laukaistakaan. Muut tekijät ovat yleensä tarpeen laukaisuun.
- Tietyt kasvihormonit , kuten gibberelliinit , ovat tärkeitä kukinnan stimuloinnissa pitkän päivän olosuhteiden puuttuessa, mutta vaikutusmekanismi on edelleen tuntematon.
Kukkien muodostuminen
Kun heterokroninen geeni on muuttanut meristeemin kasvullisesta generatiiviseksi meristeemiksi, todellinen kukkien muodostuminen tapahtuu suuren määrän vuorovaikutuksessa olevien säätelygeenien kautta.
Symmetriageenit
Niin kutsutut symmetria- tai kadastrigeenit määräävät kukan rakenteen ja sen symmetriatasot sekä kukkien ja kukkaelinten lukumäärän. Näiden geenien mutaatiot muuttavat kukan arkkitehtuuria, geenit on usein nimetty tällaisten mutaatioiden muuttamien fenotyyppien mukaan : Cycloidea (CYC) - tai Dichotoma (DICH) -geenien mutaatiot snapdragonissa tuottavat säteittäisiä symmetriakukkia tavallisten tsygomorfisten kukkien sijaan. Nämä mutaatiot esiintyvät myös luonnossa ja niitä kutsutaan peloriksi . Kukkakaali (CAL) -geenin mutaatio Arabidopsiksessa ja parsakaalissa tuottaa monia haaroja kukinnassa ilman toiminnallisia kukkia. CAL estää TFL1 -geeniä, joka edistää haarautumista kukinnassa ja on versojen identiteettigeeni. Näin ollen tfl1-mutantit muodostavat Arabidopsiksessa vähän tai yksikukkaisia kukintoja .
Kukka -identiteettigeenit

Tuloksena olevan haarautuvan kukinnan sisällä meristeemistä tulee kukkameristeemejä, kun Leafy (LFY) - ja Apetala1 (AP1) -geenit ilmentyvät . Molemmat koodittavat transkriptiotekijöitä ja kuuluvat kukkasignaalien integroiviin geeneihin. Jonka toimivuutta ja useimpien muiden geenien saatiin Arabidopsis , leijonankidat ja petunia hybridit . Tärkeimmät integrointigeenit ovat:
- FLC (Flowering Locus C) yhdistää vernalisointisignaalit ja itsenäiset signaalit. FLC on transkriptiotekijä, sen eliminaatio johtaa varhaiseen kukintaan. FLC -proteiinin / mRNA: n määrä korreloi kukinnan ajan kanssa. FLC tukahduttaa kukinnan aktivaattorin SOC1.
- SOC1 (vakioiden yliekspression vaimennin) aktivoidaan gibberelliinien ja CO: n kautta Langtagin avulla ja tukahdutetaan FLC: llä. Repression poistaminen FLC: llä ei riitä SOC1: n aktivoitumiseen: CO: n tai gibberelliinien on aktivoitava.
- Kuten SOC1, LFY: n aktivoivat gibberelliinit ja Langtag. Jälkimmäinen tapahtuu SOC1: n tai Agamous-Like24: n (AGL24) kautta.
- FT (Flowering Locus T) aktivoituu pitkiä päiviä CO: n kautta ja tukahduttaa FLC: llä. FT puolestaan aktivoi AP1: n.
Kaksi viimeistä geeniä sekvenssissä ovat LFY ja AP1, niin kutsutut kukka-meristeemi-identiteettigeenit, mutta ne ovat vuorovaikutuksessa keskenään: AP1 aktivoidaan LFY: llä; kuitenkin kukka muodostuu myös ilman LFY: tä, mutta ei oikeilla elimillä. Jo mainittu TFL1 ja toisaalta LFY ja AP1 tukahduttavat toisiaan. TFL1: tä käytetään estämään ennenaikaista kukintaa.
Kukkaelinten perustaminen

Kukkaelimet määritetään ABC -mallin mukaan. Nämä ovat kolme geeniluokkaa, jotka vuorovaikutuksen kautta määrittävät kukkaelinten identiteetin (elinidentiteettigeenit):
- A johtaa sepals
- A ja B johtavat terälehtiin
- B ja C johtavat heteisiin
- C johtaa karppiin
A ja C tukahduttavat toisiaan niin, että päällekkäisyyksiä ei ole. Jos yhtä ei ole, toinen luokka muodostuu koko kukkaan. ABC -mallin perusti E. Coen ja E. Meyerowitz vuonna 1991 , ja se on sittemmin vahvistettu ja laajennettu useita kertoja.
Niin pitkälle, seuraavat geenit tunnetaan in Arabidopsis :
- Luokka A: AP1 ja AP2
- Luokka B: AP3 ja Pistillata (PI)
- Luokka C: Agamous (AG)
Nämä geenit ilmentyvät jatkuvasti niitä määrittävissä elimissä, ei vain alussa. AP1 ei ole vain kukka -meristeemigeeni, vaan myös elinidentiteettigeeni. Kukkien muodostumisen alkuvaiheessa se muodostuu koko meristeemiin, mutta myöhemmin vain kukkakuoren ympyröihin. Mutaatiot johtavat elinten identiteetin muutokseen. Jos A epäonnistuu, karvat kehittyvät lehtien sijaan ja heteet terälehtien sijasta (koska C on nyt muodostunut). Kolmoismutantit muodostavat vain normaaleja lehtiä. C -mutanteilla ei ole rajoitettua kasvua, meristeemi jatkaa kasvuaan loputtomiin.
Lisäksi löydettiin neljäs luokka, E, joka Arabidopsiksessa koostuu neljästä geenistä Sepallata 1-4 (SEP). Ne ilmaistaan terälehdissä, hedeissä ja karpeissa ja ovat välttämättömiä elinten identiteetin kannalta. Kolmoismutantit (1-3 jaksot) muodostavat vain lehtilehtiä, nelinkertaiset mutantit vain lehtiä.
Viides luokka, D, koostuu geeneistä, jotka ovat vastuussa munasolujen identiteetistä . In Petunia nämä ovat geenit, kukat Binding Protein (FBP) 7 ja 11. Kaksoismutatit muodostavat emi-rakenteita sijasta ovules. In Arabidopsis , vastaavat geenit ovat Seedstick (STK), Shatterproof1 (SHP1) ja SHP2.
Suurin osa kukka -elingeeneistä kuuluu MADS -laatikkogeenien perheeseen . MADS -laatikkogeenit ovat laajalti levinneet kasveihin. Niitä löytyy muun muassa kasvien homeoottisista geeneistä (kuten AGAMOUS ja DEFICIENS), jotka osallistuvat kasvien elinten identiteetin kehittämiseen. Esimerkki tästä on kukkaelinten määritelmä. ABCE -geenit ovat välttämättömiä, mutta myös riittäviä kasvuselinten kukkaelinten kehittymiselle. Tämä voidaan osoittaa geenien ektooppisen ilmentymisen avulla.
Kuinka kukkien identiteettigeenit, jotka ilmentyvät koko kukka -meristeemissä, hallitsevat elinidentiteettigeenejä, ovat vasta alkaneet selvittää. Ensinnäkin B-luokan geeni AP3 näyttää ilmentyvän LFY: n, AP1: n ja epätavallisten kukkaelinten (UFO) geenin yhdistelmän kautta. LFY aktivoi AG: n, C-luokan geenin, ilmentymisen toisen koaktivaattorin, Wuschelin (WUS) kanssa. Aktivoinnin jälkeen AG tukahduttaa WUS: n uudelleen. Tämä rajoittaa kukka -akselin kasvua.
Theißenin kvartettimalli vuonna 2001 toi uuden askeleen ymmärrykseen. Siinä sanotaan, että kukkaorganisaation proteiinit toimivat tetrameereissä (siis kvartetti). Siksi jokaiselle elimelle on olemassa ainakin yksi erityinen tetrameeri. Ainakin hiivassa on jo osoitettu, että proteiinit todella muodostavat tetrameerejä. Malli toimii jo esimerkkinä monille töille.
ABC -mallin monimuotoisuus

Vertailututkimukset useista kasvilajeista ovat osoittaneet, että edellä kuvatussa muodossa olevaa ABC -mallia tulisi soveltaa vain eudicotyledoneihin . On olemassa erilaisia ominaisuuksia, erityisesti angiospermien perusryhmissä. In lampi ruusut ( Nuphar ) ei ole teräviä rajoja geenin ilmentymistä ( ”fading rajojen”), niin että on olemassa siirtymä sarja calyx kaltainen tepals kautta aateliskruunu kaltainen tepals on staminodes ja heteet ja carpels.
Tulppaani muodostaa yhtenäisen kukka kirjekuori kaksi ympyrää. Täällä luokan B geenit ilmentyvät molemmissa piireissä.
Kukkia lännenmaarianheinä on hyvin vahvasti johdettu. Ne osoittavat kuitenkin ABC -mallin, jonka mukaan palea on merkitty A: lla ja kavernoidut kappaleet (lodiculae) A: lla ja B: llä, eli homologinen verhiön ja kruunun suhteen.
Telakka ( Rumex ) muodostaa yhtenäinen, vaan vähentää ja huomaamaton kukka kansi. Täällä B -luokka rajoittuu heteisiin, joten molemmat kirjekuoren ympyrät määritellään vain A.
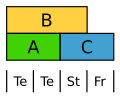
Tulppaanin muokattu ABC -malli. Te Teals
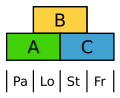
ABC -malli riisille. Pa palea, katso erektiokudosta
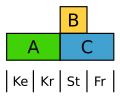
Muutettu telakan ABC -malli



Sillä välin A-luokan yleinen pätevyys kukka-elinidentiteettigeeneinä on kuitenkin kyseenalaistettu. Kuten edellä todettiin, luokan A geeneillä on myös rooli kukkien meristeemin identiteetin määrittämisessä. Toistaiseksi ne voidaan todistaa vain Arabidopsiksen kukkaelinten muodostumisessa, ja niiden pitäisi olla tärkeitä vain lähisukulaisille. In Antirrhinum , BC geenit ovat riittävät kukan muodostumista. Missä määrin tämä koskee muita angiosperm -ryhmiä, on avoin.
Teesi
Kukintaprosessia kutsutaan antesiaksi, kukkaelinten kehitysvaiheksi silmutilojen lopusta tai silmujen kehityksen alusta kuihtumisen alkuun.
Kukka -aukko
Ennen avaamista, kukka muodostaa alkuunsa samanlainen lepotilatiedosto nuppuja. Terälehdet toimivat suojana. Kukkien avaaminen on yksi kasvien liikkeistä . Kukannupun avaaminen voi tapahtua eri mekanismien avulla:
- Käänteinen ionien kertyminen. Toistaiseksi tämä on osoitettu selvästi vain Kochin gentian ( Gentiana acaulis ) tapauksessa.
- Ohjelmoitu solukuolema terälehtien määritetyillä alueilla.
- Veden menetys päivällä ja täydentäminen yöllä. Tämä tapahtuu esimerkiksi Silene saxifragan kanssa , joka kukkii yöllä ja jonka terälehdet käpristyvät päivällä veden menetyksen vuoksi.
- Terälehtien ulko- ja sisäpuolen erilainen kasvu. Siinä tapauksessa tulppaanin , sisällä terälehdet on kasvun optimi, joka on 10 ° C alempi kuin sen ulkopuolella, joka johtaa aukkoon aamulla ja sulkeminen illalla.
Tärkeä tekijä terälehtien leviämiselle on turgorin lisääntyminen , mikä johtuu yleensä sokeripitoisuuden noususta korkeamolekyylisien hiilihydraattien ( esimerkiksi ruusujen tärkkelys , fruktaanit ja päivänliljat ) hajoamisen seurauksena. . Tähän liittyy soluseinän laajentuminen. Kasvihormonien roolia koskevat tiedot ovat kirjallisuudessa ristiriitaisia.
Kukkien avautuminen johtuu ulkoisista tekijöistä. Yöllä kukkivilla lajeilla liiallinen kosteus aiheuttaa usein illalla. Varhaisen kevään kukkijat reagoivat erityisesti lämpötilan nousuun , esimerkiksi lumikello ( Galanthus nivalis ) tai krookukset ( Crocus spp.). Kolmas ryhmä reagoi valoon, kuten päivänkakkara ( Bellis perennis ). Pitkäikäisten kukkien avaaminen ja sulkeminen päivällä ja yöllä tapahtuu samojen mekanismien kautta ja on altis endogeeniselle rytmille. Kukkien avaamisen ja sulkemisen solujen fysiologista ja erityisesti geneettistä taustaa ei tunneta toistaiseksi.
Kukkien sulkeminen voi tapahtua erilaisen kasvun tai palautuvien turgorimuutosten kautta. Näissä tapauksissa toistuva avaaminen ja sulkeminen on mahdollista. Vanhenemisen aiheuttama turgorin menetys johtaa kukan pysyvään sulkemiseen.
pölytys

Pölytys on urospuolisen siitepölyn siirtymistä hedelmöittymisen naispuolisiin elimiin: mikrotyyppiin voimisteluissa, arpia angiospermiin. Pölytys ei siis ole sama kuin lannoitus. Pölytys voi tapahtua saman henkilön siitepölyllä ( itsepölytys , autogamia) tai toisen yksilön siitepölyllä ( ristipölytys , allogamia).
Kuitenkin itsepölytys vähentää geneettistä vaihtelua. Kasveissa on useita mukautuksia itsepölytyksen tai -lannoituksen välttämiseksi:
- Herogamia on porkkanan ja häpeän alueellinen erottaminen niin, että itsepölytys ei ole mahdollista. Vuonna caper holkki ( Capparis spinosa ), koko emi nostetaan avulla gynophor, niin että arpi on edellä ponsien lähestymistavassa polku pölyttäville hyönteisille.
- Dichogamy on heteiden tai munasarjojen kypsyyden ajallinen erottaminen. Näin ollen on olemassa urospuolisia ( proterandry ) ja naaraspuolisia kukkia ( proterogyny ). Samanaikaista kypsyyttä kutsutaan homogamiaksi.
Dichogamy ja hercogamy voivat estää pölytyksen kukassa, mutta eivät yhdestä kukasta toiseen saman kasvin (geitonogamia). Siksi monet lajit ovat kehittäneet lisää välttelymekanismeja:


- Itse yhteensopimattomuus : geneettiset tekijät estävät itsensä hedelmöittymisen. Tällainen ristiriita järjestelmät ovat usein myös morfologisesti tunnistettavissa (heteromorphism): Tunnettu esimerkki on heterostyly on primroses ( primula ).
Pölyttäjän tyypistä riippuen on kolme suurta sopeutumisoireyhtymää: tuulenpölytys (anemofilia), vesi (hydrofiilisyys) ja eläimet (zoophilia). Voimistelijat ovat ensisijaisia tuulipölyttäjiä, kun taas ensimmäiset selkärankaiset olivat todennäköisesti pääasiassa eläinten pölyttämiä. Tuuli- ja vesipölytys kehittyivät vain toissijaisesti angiospermien sisällä. Kunkin oireyhtymän pääpiirteet ovat:
- Anemofilia : huomaamattomat kukat; vähentynyt elinten määrä; Mono- tai diocyte usein; tiheät, usein riippuvat kukinnot; vähän tai ei lainkaan siitepölyä; sileä siitepölypinta; Suuret pinta -arvet; yksi tai muutama munasolu kukkaa kohti; ei mesiä.
- Hydrofiilisyys : huomaamattomat kukat; Mono- tai diocyte usein; Ilmakudosten ulkonäkö; kostuttamattomat siitepölyseinät; langan kaltaiset siitepölyjyvät; Suuret pinta -arvet; yksi tai muutama munasolu kukkaa kohden.
- Zoofilia : Pölytettyjen kasvien tapauksessa ei keskity kukkaan morfologisena yksikkönä, vaan kukka toiminnallisena yksikkönä. Kukka vastaa usein kukkaa (tulppaani), mutta usein monet kukat yhdistetään yhdeksi kukkaksi , jota sitten kutsutaan pseudanthiumiksi . Esimerkkejä ovat kaikki päivänkakkara perhe (kuten päivänkakkara ) ja umbelliferae ( porkkana ). On harvinaisempaa, että yksi kukka muodostaa useita kukkia ( meranthium ), kuten iiriksen tapauksessa . Zoofiilisten kukkien pääominaisuudet ovat: hermafrodiittikukat tai pseudanthia; Angiospermia; räikeä väri; voimakas tuoksu; Siitepöly tai mesi elintarvikkeena tai petosvälineet; voimakkaasti veistetty siitepölypinta ja paljon siitepölyä.
lannoitus
Nacktsamernissa siitepölyjyvät pääsevät munasolujen mikrotyyliin. Yleensä ne imeytyvät siitepölykammioon kuivaamalla pölytyspisarat. Siitepölykammiossa klaanista riippuen siittiöitä vapautuu tai siitepölyputket itävät. Pölytyksen ja lannoituksen välillä voi kulua jopa kuusi kuukautta (jotkut sykliset).
Angiospermissä siitepölyjyvät pääsevät emän leimautumiseen. Siitepölyn putki kasvaa läpi kynää erityinen johtavan siiteputki ja näin ulottuu leimautumisen että ovuleina munasarjoissa. Jos tyylissä on vain yksi yhtenäinen kudos, niin että siitepölyputket voivat päästä yhden kappelin leimautumisesta toisen kappelin munasoluihin, johtavaa kudosta kutsutaan kokonaisuudeksi compitum .
Kun siitepölyputket tai siittiöt ovat saavuttaneet munasolut, todellinen hedelmöitys tapahtuu. Kanssa koppisiemenisiä, gnetum ja efedraa on kaksinkertainen lannoitus: kanssa koppisiemenisistä toinen siittiöiden ytimet sulakkeita munasolun ja muodostaa tsygootti . Toinen sulake jo diploidiseen alkionpussin ytimeen muodostaa triploidisen endospermin ytimen, josta siementen ravitseva kudos ( endospermi ) syntyy. Tapauksessa efedra, toisen siittiöiden ydin sulakkeita vatsan kanava solun archegonium, tapauksessa gnetum, kaksi siittiösolujen fuusioituvat kaksi gametofyytin soluja. Yleensä vain yksi kahdesta tuotetusta zygootista kehittyy.
Hedelmöityksen jälkeen tsygootista kehittyy alkio, munasolusta siemen ja kukasta hedelmä.
Vanheneminen
Kasveissa sukupuolieliminä kukkia muodostetaan jatkuvasti uudelleen, toisin kuin eläimillä. Elinikä mitataan tarkasti, koska kukat kuluttavat suuria resursseja ja leima on myös tärkeä taudinaiheuttajien saapumispaikka. Myös jo pölytetyt kukat kilpailevat tarpeettomasti pölyttäjistä niiden kanssa, joita ei pölytetty.
Siitepölyllä pölyttäminen on tärkeä ikääntymisen (ikääntymisen) laukaisija . Useimmissa lajeissa tämä lyhentää huomattavasti kukan elinikää. Monissa kasveissa pölytys laukaisee kasvihormonin eteenin muodostumisen , mikä puolestaan laukaisee terälehtien vanhenemisen. Muut lajit ovat kuitenkin herkkiä eteenille ja niiden vanhenemismekanismi on tuntematon. Elinten tasolla kruunu, heteet ja stylus kuolevat pölytyksen jälkeen, kun taas munasarja kehittyy hedelmäksi. Kuolema kulkee käsi kädessä ainesosien uudelleensijoittamisen kanssa, joka muistuttaa lehtien vanhenemista.
evoluutio
Angiospermin kukinnan kehitys ei ole selvää, koska "modernin" angiospermin kukinnan fossiilisia esiasteita on tuskin olemassa. Sienien ja muiden siemenkasvien väliset suhteet ovat myös edelleen epäselviä. Kukkien alkuperästä on kaksi erilaista hypoteesia. Syynä hermaphrodite angiosperm -kukinnan muodostumiseen oletetaan olevan sopeutumista hyönteisten (kovakuoriaisten) pölyttämiseen; munasolujen siirtyminen suljetuiksi karpeiksi suojaisi siten kovakuoriaisten puremista.
Arberin ja Parkinin Euanthienheorie (1907) olettaa, että angiospermien esi-isillä oli jo hermafrodiittisia kukkia ja siksi angiosperm-kukka on yksiakselinen järjestelmä, jossa on sivuttaiset mikro- ja megasporofyllit. Heteet ja karvat ovat siksi homologisia lehtiin nähden. Tämä teoria perustuu analyyseihin fossiilisesta suvusta Caytonia , jonka uskotaan olevan angiospermien mahdollinen esi -isä. Caytonialla oli pinnate mikro- ja megasporofylli, jotka ehkä seisoivat hermafrodiittisissa kukissa. Megasporofyllissä kupit, joissa jokaisessa oli useita munasoluja, istuivat rachiksen puolella . Mikrosporofyllit olivat myös pinnate, jokaisessa höyhenessä oli useita ryhmiä umpeen kasvaneita siitepölypusseja. Karvas angiospermit olisi voitu luoda, kun rachis olisi tasainen, hede pelkistämällä varreksi, jossa on kaksi synangiaa, joissa molemmissa on kaksi siitepölypussia.
Richard Wettsteinin teoria pseudanthiasta olettaa, että angiosperm -kukka syntyi unisexual kukkien kukinnasta . Niinpä heteet ja karpeet olisivat sivuvaroja eivätkä lehtiä. Tämä teoria perustuu oletukseen, että angiospermit ovat peräisin Gnetopsidasta. Kappeli luotiin kukan kantaosasta, munasolun toisesta osasta kukan alla olevasta luukusta. Tätä teoriaa tuskin tukevat molekyyli- ja morfologiset analyysit.
Vaikka useimmat kirjoittajat suuntaavat nyt euanthia -teoriaan, heteen, kappelin ja munasolujen toisen osan alkuperä on edelleen epäselvä.
Molekyylibiologiaan perustuvat teoriat
Vanhimmat angiosperm -kukat olivat hermafrodiittisia ja niillä oli eriytymätön periantti spiraaliasennossa tai enemmän kuin kaksi pyörteitä.
Siemenkasvien ulkopuolella ei ole kukka -elinidentiteettigeenejä tai niiden ortologeja . Voimisteluissa, jotka muodostavat vain unisexual kukkia, on luokkien B ja C ortologit. Niiden ilmentyminen on samanlainen kuin angiospermsissä: C muodostuu kaikissa lisääntymiselimissä, B uroskukissa. Näiden havaintojen perusteella esitettiin useita hypoteeseja hermaphroditic angiosperm -kukinnan alkuperästä:
- Theissen et ai. Vuonna 2002 urospuoliset kartiot muodostivat naaraselimiä vähentämällä B-luokan ekspressiota ylemmällä kartioalueella. "Naisen ulkopuolella" -hypoteesin mukaan urospuoliset elimet muodostuivat naaraskartioiden alapäähän. Näiden kahden mallin mukaan kukinto syntyi vasta hermafrodiitin jälkeen.
- "Enimmäkseen miespuolinen" hypoteesi perustuu myös uroskukkiin, ensimmäisenä Frohlichin ja Parker 2000: n ehdottamana. He havaitsivat, että Leafy -geeni esiintyy kahdessa kopiossa gymnospermsissä (paralogueissa). Leafy määrittää urospuoliset, tarpeelliset naaraskukat. Tarpeenmukaista esiintyy kaikissa voimisteluissa , paitsi Gnetumissa , mutta ei angiospermissä. Teoria sanoo nyt, että deaktivoimalla Needly -geenin uroskartioista tuli hermafrodiittisia.
Angiospermin kukinnan kehitys tapahtui neljässä keskeisessä tapahtumassa:
- Hermafrodiitin kukka -akselin kehitys
- Puristetun kukka-akselin kehitys ja kasvun rajoittaminen: Tämän tekivät C-luokan geenit, jotka tukahduttavat Wuschelin , meristeemin ylläpitogeenin.
- Petaloid -kukka -kirjekuoren kehitys
- Klassisen kahden ympyrän, kaksikomponenttisen kehityksen kehittyminen verhiöstä ja kruunusta.
Soltis et ai. (2007) keskustelevat hypoteesista, jonka mukaan selkärankojen pohjalla ei ollut ABC -mallia kuten Arabidopsiksessa , vaan järjestelmä, jossa on sumeat reunat, kuten lammen ruusussa. Tästä perusmallista ABC -malli ja muokatut muodot, kuten tulppaani, voivat sitten yksinkertaisesti syntyä rajojen erilaisella teroittamisella.
Ihmisten käyttö


Toisin kuin kukista nousevat hedelmät, kukilla on toissijainen rooli ihmisten ravintokasveina. Kukkakaalin ja artisokan kukinnot toimivat vihanneksina, Cannabis sativa subsp. indica päihteenä. Joitakin kukkia tai kukintoja käytetään salaattina tai sen koristeena ( orvokit , koiranputkea , nasturtiumia ). Kukat, silmut, kukkaosat tai kukinnot tarjoavat mausteita: Mugwort ( Artemisia vulgaris subsp. Vulgaris ), laventeli ( Lavandula angustifolia ), neilikka ( Syzygium aromaticum ), sahrami ( Crocus sativus ) ja humala ( Humulus lupulus ). Monia kukkia käytetään tuoksuöljyjen , kuten laventeliöljyn ja ruusuöljyn, valmistukseen .
Kukilla on kuitenkin paljon suurempi rooli koristeina koristekasvien ja leikkokukkien muodossa . Maailman suurimpana leikkokukkien tuojana Saksa toi vuonna 2004 tavaroita, joiden tukkuarvo oli 1,1 miljardia euroa. Leikkokukkien ja ruukkukasvien maailmanmarkkinat vuonna 2000 olivat 6,8 miljardia euroa.
Aamunkoiton jumalatar ( Aurora / Eos ) hajottaa kukkia uuden päivän valonsäteiden symbolina. Kukat ovat myös jumalatar Floran ominaisuus . Neljän vuodenajan tapauksessa kukat on määrätty kevääseen ja viiden aistin tapauksessa hajuaistiin . Kukat ovat olleet merkki kauneuden ja elämän ohimenevyydestä muinaisista ajoista lähtien. Asetelmissa kuihtunut kukka tai pudonneet terälehdet näkyvät aina vanitas -motiivina. Logiikan ja toivon allegorioita kuvataan joskus kukilla käsissään, mikä symboloi, että kukka kehittyy pian hedelmäksi. Yksittäisten lajien on omat symbolisia merkityksiä, joka voi muuttaa vuosisatojen. Ruusu oli ja on määrite on Venus , Dionysoksen ja Mary , The symboli jumalallisen ja maallisesta rakkaudesta , sillä neitsyet vaan myös prostituutioon .
Vuonna kuvataide , kukkia ja kukkia kukka-aiheita ovat usein esitetään teema, lähinnä ilmaista tuoreuden ja iloa.
Alaviitteet
- ↑ "kukka määrätyllä, puristetulla, biseksuaalisella lisääntymisakselilla, joka koostuu megasporangiasta (karselit), mikrosporangiasta (heteet) ja steriilistä periantista, joka koostuu vähintään yhdestä steriilistä laminaarisesta elimestä". G. Theissen, R. Melzer: taustalla olevia molekyylitason mekanismeja alkuperä ja monipuolistaminen Angiosperm Flower . Julkaisussa: Annals of Botany. Vuosikerta 100, 2007, s.603-619.
- ↑ hyvin samanlainen kuin määritelmä, joka perustuu kirjallisuuskatsaukseen julkaisuissa: Richard M.Bateman, Jason Hilton, Paula J.Rudall: Angiospermien morfologinen ja molekyylifylogeneettinen konteksti: käytettyjen ylhäältä alas- ja alhaalta ylös -lähestymistapojen vastakohtana päätellä ensimmäisten kukkien todennäköiset ominaisuudet . Julkaisussa: Journal of Experimental Botany. Vuosikerta 57, 2006, s. 3471-3503, doi : 10.1093 / jxb / erl128 .
- ↑ katso Wouter G. van Doornin arvostelu, Uulke van Meeteren: Kukkien avaaminen ja sulkeminen: arvostelu. 2003.
- ↑ Osio perustuu: Wouter G. van Doorn, Uulke van Meeteren: Kukkien avaaminen ja sulkeminen: arvostelu. 2003.
lähteet
Artikkeli perustuu pääasiassa seuraaviin lähteisiin:
- Peter Leins, Claudia Erbar: Kukka ja hedelmät. Morfologian, evoluutiohistorian, fylogenian, toiminnan ja ekologian näkökohdat . Schweizerbart, Stuttgart 2000, ISBN 3-510-65194-4 .
- Peter Sitte , Elmar Weiler , Joachim W.Kadereit , Andreas Bresinsky , Christian Körner : Kasvitieteiden oppikirja yliopistoille . Perustaja Eduard Strasburger . 35. painos. Spectrum Academic Publishing House, Heidelberg 2002, ISBN 3-8274-1010-X .
Kukkien induktio- ja kukkienmuodostusosuudet perustuvat:
- Peter Schopfer, Axel Brennicke: Kasvien fysiologia . Perusti Hans Mohr . 6. painos. Elsevier, Spektrum, München / Heidelberg 2006, ISBN 3-8274-1561-6 , s. 501-523 ( rajoitettu esikatselu Google-teoshaussa).
- Thomas Jack: Molekyyli- ja geneettiset mekanismit kukkakontrollista . Julkaisussa: The Plant Cell. Osa 16, täydennys 1, 2004, s. S1-S17, doi : 10.1105 / tpc.017038 .
- Paul K.Boss, Ruth M.Bastow, Joshua S.Mylne, Caroline Dean: Useita polkuja kukinnan päätöksessä: käyttöönotto, edistäminen ja nollaus. Julkaisussa: The Plant Cell. Volume 16, Supplement 1, 2004, s.S18 -S31, doi : 10.1105 / tpc.015958 .
Muuta tärkeää kirjallisuutta:
- Günter Theißen, Rainer Melzer: Molekyylimekanismit, joiden taustalla on angiospermikukka . Julkaisussa: Annals of Botany. Osa 100, nro 3, 2007, s. 603-619, doi : 10.1093 / aob / mcm143 .
- Wouter G. van Doorn, Uulke van Meeteren: Kukkien avaaminen ja sulkeminen: arvostelu. Julkaisussa: Journal of Experimental Botany. Osa 54, nro 389, 2003, s. 1801-1812, doi : 10.1093 / jxb / erg213 (kukka-aukko).
Yksilöllisiä todisteita
- ^ Friedrich Kluge , Alfred Götze : Saksan kielen etymologinen sanakirja . 20. painos. Toimittanut Walther Mitzka . De Gruyter, Berliini / New York 1967; Uusintapainos (”21. muuttumaton painos”) ibid 1975, ISBN 3-11-005709-3 , s.87 .
- ↑ a b Gerhard Wagenitz : Kasvitieteellinen sanakirja. Termit historiallisessa kontekstissaan . Toinen laajennettu painos. Spectrum Academic Publishing House, Heidelberg / Berliini 2003, ISBN 3-8274-1398-2 , s. 47 f .
- ↑ Peter Schütt , Hans Joachim Schuck, Bernd Francisco (toim.): Puu- ja pensaslajien sanasto . Metsätieteiden vakiotyö. Tärkeiden puu- ja pensaslajien morfologia, patologia, ekologia ja järjestelmällisyys . Nikol, Hampuri 2002, ISBN 3-933203-53-8 , s. 83 (uusintapainos 1992).
- ^ Manfred A.Fischer, Wolfgang Adler, Karl Oswald: Retkikasvillisuus Itävaltaan, Liechtensteiniin ja Etelä -Tiroliin . Toinen, parannettu ja laajennettu painos. Ylä-Itävallan maakunta, Ylä-Itävallan osavaltion museoiden biologiakeskus, Linz 2005, ISBN 3-85474-140-5 , s. 90 .
- ^ Rudolf Schubert , Klaus Werner, Hermann Meusel (toim.): Exkursionsflora for the DDR and FRG . Perustaja: Werner Rothmaler. 13. painos. nauha 2 : verisuonikasvit . Ihmiset ja tieto, Berliini 1987, ISBN 3-06-012539-2 , s. 24 .
- ^ Siegmund Seybold (toim.): Schmeil-Fitschen interaktiivinen . CD-ROM, versio 1.1. Quelle & Meyer, Wiebelsheim 2002, ISBN 3-494-01327-6 .
- ^ Peter Sitte , Elmar Weiler , Joachim W.Kadereit , Andreas Bresinsky , Christian Körner : Kasvitieteiden oppikirja yliopistoille . Perustaja Eduard Strasburger . 35. painos. Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X , s. 750 .
- ↑ Herderin biologian sanakirja . CD-ROM. Spektrum Akademischer Verlag, Heidelberg 2003, ISBN 3-8274-0354-5 , avainsana "Blossom".
- ^ Peter Sitte , Elmar Weiler , Joachim W.Kadereit , Andreas Bresinsky , Christian Körner : Kasvitieteiden oppikirja yliopistoille . Perustaja Eduard Strasburger . 35. painos. Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X , s. 720, 723 .
- ^ Katja E. Jaeger, Alexander Graf, Philip A. Wigge: Kukinnan hallinta ajassa ja avaruudessa. Julkaisussa: Journal of Experimental Botany. Osa 57, nro 13, 2006, s. 3415-3418, doi : 10.1093 / jxb / erl159 .
- ↑ Laurent Corbesier, George Coupland: Florigen -etsintä: katsaus viimeaikaiseen edistymiseen. Julkaisussa: Journal of Experimental Botany. Osa 57, nro 13, 2006, s. 3395-3403, doi : 10.1093 / jxb / erl095 .
- ↑ Enrico S.Coen, Elliott M.Meyerowitz: Soturien sota: kukkien kehitystä ohjaavat geneettiset vuorovaikutukset. Julkaisussa: Nature. Vuosikerta 353, 1991, s. 31-37, DOI: 10.1038 / 353031a0 .
- ↑ a b c d e f g h i Günter Theißen, Rainer Melzer: Molekyylimekanismit, jotka takaavat angiospermikukan alkuperän ja monipuolistumisen. Julkaisussa: Annals of Botany. Osa 100, nro 3, 2007, s. 603-619, doi : 10.1093 / aob / mcm143 .
- ^ A b Douglas E.Soltis, André S.Chanderbali, Sangtae Kim, Matyas Buzgo, Pamela S.Soltis: ABC -malli ja sen soveltuvuus perusperäkkäisiin siittiöihin . Julkaisussa: Annals of Botany. Osa 100, nro 2, 2007, s. 155-163, doi : 10.1093 / aob / mcm117 .
- ↑ Video avata kukkia päälle YouTubessa .
- ↑ Hilary J.Rogers: Ohjelmoitu solukuolema kukka -elimissä: miten ja miksi kukat kuolevat? Julkaisussa: Annals of Botany. Osa 97, nro 3, 2006, s. 309-315, doi : 10.1093 / aob / mcj051 .
- ^ EA Newell Arber, John Parkin: Angiospermien alkuperästä. Julkaisussa: Journal of the Linnean Society of London, Botany. Osa 38, nro 1, 1907, s. 29-80, DOI: 10.1111 / j.1095-8339.1907.tb01074.x ( digitaalinen kopio ).
- ↑ Richard Wettstein: Käsikirja systemaattisesta kasvitieteestä. Osa 2, F.Deuticke, Wien 1907.
- ↑ Michael W. Frohlich, David S. Parker: Useimmiten miesten teoria kukkien evoluution alkuperästä: geeneistä fossiileihin. Julkaisussa: Systematic Botany. Vuosikerta 25, nro 2, 2000, s. 155-170, DOI: 10.2307 / 2666635 ( PDF-tiedosto ).
- ↑ a b Richard M.Bateman, Jason Hilton, Paula J.Rudall: Angiospermien morfologinen ja molekyylifylogeneettinen konteksti: vastakkain ylhäältä alas ja alhaalta ylös -menetelmien kanssa, joita käytetään päättämään ensimmäisten kukkien todennäköisistä ominaisuuksista. Julkaisussa: Journal of Experimental Botany. Osa 57, nro 13, 2006, s. 3471-3503, doi : 10.1093 / jxb / erl128 .
- ^ Wolfgang Franke : Nutzpflanzenkunde. Käyttökelpoiset viljelykasvit lauhkeilla leveysasteilla, subtrooppisilla ja tropiikilla . 4. tarkistettu ja laajennettu painos. Thieme, Stuttgart / New York 1989, ISBN 3-13-530404-3 .
- ↑ florist.de ( Memento marraskuusta 5, 2007 Internet Archive )
- ↑ ianywhere.com, Onnistunut kukkamarkkinoilla mobiilitekniikan avulla , käytetty 12. joulukuuta 2007
- ↑ Lucia Impelluso: Luonto ja sen symbolit. Kasveja, eläimiä ja myyttisiä olentoja . Parthas Verlag, Berliini 2005, ISBN 3-936324-03-4 .
- ↑ Marianne Beuchert : Kasvien symboliikka . Insel Verlag, Frankfurt / Leipzig 2004, ISBN 3-458-34694-5 , s. 279 .
jatkokäsittelyä
- Dieter Hess: Kukka. Johdanto kukkien rakenteeseen ja toimintaan, ekologiaan ja kehitykseen sekä ohjeet yksinkertaisiin kokeisiin . Toinen parannettu ja täydennetty painos. Eugen Ulmer, Stuttgart (Hohenheim) 1990, ISBN 3-8001-6434-5 (yleisesti ymmärrettävä johdanto myös maallikolle, painettu).
- Erityisnumero: Kukkivatutkimuksen pääteemat. Julkaisussa: Journal of Experimental Botany. Osa 57, nro 13, 2006 ( verkossa ).
- Douglas E. Soltis, James H. Leebens-Mack, Pamela S. Soltis (toim.): Developmental Genetics Flower (= Advances in Botanical Research . Volume 44 ). Academic Press, San Diego et ai. 2006, ISBN 0-12-005944-4 (englanti, verkossa ).
nettilinkit
- Kukkabiologia ( Muisto 19. heinäkuuta 2011 Internet -arkistossa ; PDF; 532 kB)
- Kukkivat kuvat Baselin yliopiston kuva -arkistosta
- Kukka- ja hedelmäbiologia
- Silmien itämisen ja kukkien muodostumisen fysiologia