
Tardigrade
| Tardigrade | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Karhukaiset Milgiene tardigradum | ||||||||||||
| Järjestelmällisyys | ||||||||||||
| ||||||||||||
| Tieteellinen nimi | ||||||||||||
| Tardigrada | ||||||||||||
| Spallanzani , 1777 | ||||||||||||
| Luokat | ||||||||||||
Karhukaiset (Tardigrada) - kutsutaan myös vesi kantaa - muodostavat eläimen kannan sisällä molting eläimet ( Ecdysozoa). Kahdeksanjalkaiset eläimet, jotka ovat yleensä alle millimetrin kokoisia, muistuttavat jonkin verran karhuja ulkonäkönsä ja kömpelön liikkumismallinsa vuoksi , mikä johti niiden nimeen saksankielisissä maissa . Sen tieteellinen nimi (yhdistelmä latinalaisesta tarduksesta 'hidas' ja gradus 'askel' ) palaa myös hitaaseen liikkeeseen.
He elävät maailmanlaajuisesti meressä, makeassa vedessä tai kosteissa elinympäristöissä maalla; ne ovat erityisen yleisiä sammal tyynyissä . Yksi eläinten ominaisuus on kryptobioosi , kuoleman kaltainen tila, jossa ne voivat selviytyä äärimmäisistä ympäristöolosuhteista. Voi karhukaiset sekä rehun sisällöstä kasvisolujen ja saalis on pieniä eläimiä, kuten sukkulamadot (Nematoda) tai rataseläimet (Rataseläimiä), jossa ne lävistävät sen ja imeä. Tardigrade lisääntyy yleensä seksuaalisesti . Jotkut lajit lisääntyvät kuitenkin myös partenogeneettisesti eli ilman urosten osallistumista; naarasmunat kehittyvät tässä tapauksessa ilman hedelmöitystä.
Seuraavana Viime omaiset karhukaiset lienevät raajat (Arthropoda) ja Velvet madot (käsnäjalkaiset), joiden kanssa he taksoniin Panarthropoda muodossa.
rakentaminen

Koko karhukaiset on välillä 50 mikrometriä joidenkin nuorten ja 1,5 mm varten lajin suvun Macrobiotus , mutta on yleensä noin 100-500 mikrometriä, niin että eläimet lasketaan kuten meiofauna . Heillä on suhteellisen täyteläinen, lieriömäinen runko, joka on litteä vatsaan ja pinnalta näyttää koostuvan neljästä kehon osasta, joista jokaisella on pari jalkaa ja pääosa. Tämä ulompi vaikutelma, joka johtuu ihon uloimman kerroksen poikittaisista taitteista, on kuitenkin harhaanjohtava: pää, joka on vain hieman erotettu ja erotettu muusta kehosta, ei luultavasti koostu yhdestä, vaan kolmesta toisiinsa liittyvästä osasta; runko ei sitä vastoin voisi koostua neljästä, vaan mahdollisesti viidestä osasta.
Merilajit ovat enimmäkseen värittömiä tai huomaamattomia valkoisesta harmaaseen, kun taas maa- ja makean veden muodoissa on usein värejä, kuten punainen, keltainen, vihreä, vaaleanpunainen, violetti tai musta. Värin aiheuttavat joko pigmentit ulommassa ihossa, kynsinauhassa tai kehon ontelon, hemocoeloman tai suolen värillinen sisältö . Värit myös usein siirtyvät suolistosta hemo- keeloomaan ja sieltä kerääntyvät kynsinauhaan.
Tyypillistä on, että monet tardigrade -kudoksista ovat ruma , eli niiden solujen lukumäärä on geneettisesti määrätty. Sen vuoksi koon kasvu ei voi tapahtua lisäämällä solujen määrää, vaan vain yksittäisten solujen kasvun kautta. Jos toinen kahdesta tytärsolusta tapetaan ensimmäisen solujakautumisen jälkeen (kokeellisella toimenpiteellä), kehittyy anatomisesti täydellinen ja elinkelpoinen eläin, joka sisältää sitten vain puolet enemmän soluja kuin normaalisti.
raajat

" Kantojalka" , rungon saumaton ulkonema, kuten myös vastaavassa stumpefoots -ryhmässä , syntyy neljästä rungon segmentistä vatsan vasemmalla ja oikealla reunalla (ventrolateral) . Joskus jalan yksittäiset alueet erotetaan toisistaan, joista käytetään nimitystä coxa ("lonkka"), reisiluu ("reisi"), sääriluu ("shin") tai tarsus ("jalka"). Koska jalat ovat vain hieman eriytyneet, näiden termien anatomiset perusteet ovat kyseenalaisia; Joka tapauksessa ne on kuitenkin ymmärrettävä puhtaasti kuvaavina, ts. Ne eivät osoita evoluutiosuhdetta niveljalkaisten vastaavien jalkakomponenttien kanssa.
Monet merilajit pystyvät teleskooppaamaan jalkansa toisiinsa; Lyhyet lihakset, jotka kulkevat kaiken tardigraden jalan sisällä, auttavat heitä tässä. Jalan päässä on yleensä neljästä kahdeksaan, harvemmin jopa kolmetoista sisäänvedettävää kynsiä, jotka joskus istuvat ohuilla jalkojen ulkonäöillä, "varpailla" ja joita erittävät erityiset rauhaset, "kynsirauhaset" . Joillakin lajeilla kynnet ovat toissijaisia liimalevyille nähden; ei ole kynnet on suvun Apodibius . Usein neljännen jalkaparin kynnet osoittavat vastakkaiseen suuntaan kuin kolmen ensimmäisen parin kynnet.
iho

Kehoa rajoittaa ulkopuolelle ei-soluinen ulkokerros, kynsinauha ja yksittäinen soluista koostuva sisäkerros, epidermis . Flagelloidut solut eivät näy tardigrade -ihossa.
Kynsinauha on 0,5 mikrometriä paksu ja koostuu kolmesta kerroksesta, ulompi epikutiikki , jonka paksuus on noin 0,1 mikrometriä, alla oleva ihonsisäinen , jonka paksuus on noin 0,2 mikrometriä, ja lopullinen ihoputki, joka on noin 0,25 mikronia. Se koostuu kitiinistä , erilaisista glykoproteiineista , polysakkarideista , mukopolysakkarideista , lipideistä ja lipoproteiineista ja sisältää myös pigmenttejä monissa maa- ja makean veden lajeissa. Usein se on koristeltu kuoppia, piikkejä tai kuoppia ja joissakin lajeissa se on karkaistu selässä (selkä), harvemmin sivussa (sivusuunnassa), hyvin harvoin myös vatsassa (vatsa) muodostaen paksuja panssarilevyjä, skleriittejä ; nämä niveltyvät sitten niveljalkaisten tapaan. Kynsinauha, jalkakynnet sekä etu- ja takaontelon vuoraus, mukaan lukien suukappaleet, jotka sisältyvät edelliseen, stilettot, on säännöllisesti nyljetty .
Epiderma koostuu Eutelianista eli lajista, joka koostuu aina samasta määrästä soluja, yksinkertaisesta solukerroksesta, jota päällyskerros erittää. Sen erottaa kehon ontelosta vain ohut solunulkoinen erotuskerros, ns.
Hemokoelooma
Kehon ontelo ei ole todellinen coelom , eli sitä ei vuorata kudos, joka voidaan jäljittää alkion mesodermiin . Kaikki elimet erotetaan siitä vain ohuella peruskalvolla. Kehon onteloa kutsutaan siksi pseudo- tai, koska se on täynnä "verta", hemocoelomaksi . Todellinen coelom löytyy vain yhden sukurauhasen (gonad) ympäriltä.
Veri on väritöntä eikä se palvele happea. Se kiertää kehon liikkeiden kautta; todellista verenkiertoa ei kuitenkaan ole, aivan kuten erilliset verisuonet tai sydän. Se on täynnä jopa 200 solua, hemosyyttejä, jotka kelluvat vapaasti tai ankkuroidaan peruskalvoon, ja niiden ensimmäinen tehtävä näyttää olevan ravinteiden varastointi; niillä voi myös olla rooli bakteerien torjunnassa.
Lihaksisto
Toisin kuin läheinen kolumbus, tardigraden lihakset eivät koskaan näy paksuissa lihaskerroksissa. Sen sijaan rungon ja raajojen lihakset koostuvat eristetyistä, ohuista pitkittäislihaksista, jotka on kiinnitetty kynsinauhaan ja jotka koostuvat vain muutamasta, joskus jopa yhdestä lihassolusta. Ne joko toimivat hemocoelia vastaan, joka toimii nesteenä täytetyn ilmapallon tavoin hydroskeletonina, tai antagonistisesti, kuten ihmisen käsivarren taivutus- ja venytyslihakset, toisiaan vastaan. Rengaslihaksia ei löydy tardigradista. Lihassolut ovat enimmäkseen sileitä, harvoin raitoja, jälkimmäinen oletettavasti alkuperäinen tila; Laajalle levinneen hypoteesin mukaan sileät lihakset kehittyivät monille lajeille vain toissijaisesti sopeutumalla maaseudun elämään.
hengitys
Lihakset tarvitsevat happea työhönsä , jonka keskimääräisen kulutuksen tardigraden mukaan arvioidaan olevan 0,05 - 0,1 kuutiometriä tunnissa ja milligrammaa kehon painoa. Koska kehon pinta -ala ja tilavuus ovat erittäin suotuisat, erillisiä hengityselimiä ei kuitenkaan tarvita eikä niitä ole saatavilla. Koko kaasunvaihto voi tapahtua yksinkertaisella diffuusiolla ihon läpi, joka kuitenkin on peitettävä ohuella vesikalvolla.
Ruoansulatuselimet
Ruoansulatuskanava koostuu pitkästä putkesta, joka kulkee kehon edestä taakse ja joka voidaan jakaa suuonteloon, kurkkuun (nielulle), ruokatorveen, keskisuoleen ja peräsuoleen. Kolme ensimmäistä osaa on ryhmitelty yhteen esisuolaksi (ruokatorveksi), koska niiden kynsinauhojen vuori irtoaa yhdessä kutikulaarisen takamahan vuorauksen kanssa jokaisen karvan kanssa.
Lihansyöjillä tai kaikkiruokaisilla lajeilla (lihansyöjät ja kaikkiruokaiset) suun aukko on yleensä kehon etuosan lopussa (terminaali), toisaalta kasveja tai orgaanista jätettä (kasvissyöjiä ja detritivoreja) ruokkivissa lajeissa, se on usein hieman etuosan ventraalisen sivun takana (ventral-subterminal). Sitä ympäröi usein 10–30 kovan, kuutiomaisen ulkoneman rengas, niin sanotut lamellit, ja joskus se istuu suukappaleella, joka voidaan teleskooppia ulos.
Sylkirauhanen avautuu viereiseen putkimaiseen suuonteloon vasemmalla ja oikealla puolella, joka paitsi erittää ruoansulatuskanavan eritteitä, myös syntetisoi tardigradille ominaiset tikat. Nämä ovat kovia, enimmäkseen suoria, mutta toisinaan myös kaarevia neuloja, jotka yleensä sijaitsevat vastaavan sylkirauhasen sisällä (ontelossa), mutta voivat työntyä suuonteloon terävillä kärjillään. Paksuuntuneisiin takapäihin kiinnittyvät pro- ja kelauslihakset voidaan venyttää tai vetää sisään. Poikittaiset stiletto -pidikkeet yhdistävät ne suuonteloon, kun taas niiden etupää lepää omissa stiletto -tuppeissaan sisäänvedettynä. Lihansyöjälajeissa ne ovat yleensä hieman voimakkaampia kuin kasvissyöjämuodoissa.
Nielut koostuvat radiaalisesti ulospäin kulkevista raidallisista epiteelin lihassoluista , jotka on erotettu kehon ontelosta tyvikalvolla . Kurkun sisäpuoli, ontelo, on vuorattu kynsinauhoilla, ja Y-muotoisen triadidisen poikkileikkauksensa ansiosta se sopii erinomaisesti alipaineen tehokkaaseen muodostamiseen, jota käytetään imemään nestemäistä ruokaa suolistoon. Tämä pumppuperiaate on kehittynyt itsenäisesti myös muissa läheisesti sukua olevissa eläimissä, kuten rotifereissa .
Lyhyt ruokatorvi, joka koostuu kuutiosoluista ja joka oletettavasti erittää limaa, yhdistyy keskisuoleen, joka koostuu yhdestä epiteelisolukerroksesta, jossa on joskus sivuttaisia ulkonemia (mikrovilliä), joiden uskotaan lisäävän pinta -alaa. Suolistoon tullessaan ruokaa ympäröi usein peritrofinen kalvo , jonka tehtävänä on suojata herkkää suolistokudosta vieraiden elinten aiheuttamilta vaurioilta. Keskivatsas erittää hydrolyyttisiä entsyymejä , imee ravinteita ruoasta, varastoi ne rasvojen tai polysakkaridien muodossa ja luultavasti myös poistaa jätteet. Monilla lajeilla on laaja bakteerikasvillisuus, joka oletettavasti elää kommensaalisesti eli vaikuttamatta sen isäntään, mutta toimii joissakin tapauksissa myös ravinnon lähteenä.
Ruokajäämät kerätään lyhyeen peräsuoleen, joka on vuorattu kynsinauhalla, mahdollisesti muokattu, ja sitten vapautetaan ulkomaailmaan vatsan peräaukon kautta, hieman keskiviivan viimeisen jalkaparin eteen (mediaalinen-ventraalinen) . Monissa lajeissa ei ainoastaan Malpighian rauhaset, joita kuvataan tarkemmin jäljempänä, avautuvat peräsuoleen, mutta myös naaraiden munanjohtimet; suoliston viimeistä osaa kutsutaan tässä tapauksessa kloaakaksi.
Erittyvät elimet
Sulattamattomat jäämät, aineenvaihduntajätteet tai epäpuhtaudet voivat vapautua ulkomaailmaan eri tavoin. Toisaalta monet epäpuhtaudet kerääntyvät ihon alle kynsinauhaan epidermiksen avulla ja sitten irtoavat sen kanssa seuraavan vuodon aikana . Lisäksi sylkirauhaset näennäisesti ottavat erittymistoiminnon verenvuodon aikana. Suolisto ei ainoastaan ime ravintoaineita, vaan myös vapauttaa epäpuhtauksia kehon ontelosta suoliston onteloon, suoliston sisäosaan.
Monilla Eutardigrada -luokkaan kuuluvilla lajeilla on myös kolme, hyvin harvoin neljä erikoiselintä, joita kutsutaan Malpighian rauhasiksi . Kaksi niistä on sivussa (sivusuunnassa), yksi suolen takana (selkä). Ne avautuvat keski- ja peräsuolen risteyksessä, ja niitä käytetään todennäköisesti typpipitoisen jätteen poistamiseen. Vaikka ne ovat rakenteeltaan samanlaisia kuin hyönteisten samannimiset rauhaset, ne eivät luultavasti ole homologisia elimiä, mikä tarkoittaa, että evoluution kannalta ne eivät palaa yhteiseen edeltäjärakenteeseen. Koska niitä ei ole alun perin pidetyissä toisen suuren luokan, heterotardigradan , merilajeissa , niitä pidetään evoluution mukautumisena elämään makeassa vedessä ja maalla.
Joillakin heterotardigradan maaseudun muodoilla on Malpighian rauhasien sijasta erityisiä vatsaan sijoitettuja elimiä, jotka avautuvat toisen ja kolmannen jalkaparin juureen ja joilla on oletettavasti erittymistoiminto. Toisaalta todellisia nefridioita , erittäin kehittyneitä erityselimiä, ei esiinny tardigradissa.
Hermosto
Karhukaiset hermosto koostuu renkaan hermot pään ympäri kulkevan etusuolen ja kaksi paria hermo johdot käynnissä taaksepäin vatsa puolella, joka muodostaa niin sanotun köysitikkaat hermoston kautta ristikytkentöjä kussakin runko segmentissä .
Etuhermon rengas, joka koostuu suun yläpuolella olevasta supraesofageaalisesta ganglionista ja yksi saman ruokatorven alapuolelta , kaksi hermosoluryhmää, jotka kulkevat ruoansulatuskanavan hermonauhojen sivun läpi, on yhdistetty toisiinsa renkaan muodostamiseksi. primitiivinen " aivot voidaan harkita". Ylempi nielun ganglion koostuu kahdesta parista aivolohkoja, jotka sijaitsevat takana ja vastapäätä takapäätä (kaudaalisesti), sisä- ja ulommasta parista sekä lisäparista, joka sijaitsee stilettojen sivulla. Ulompi takapari, jos sellainen on, hermottaa silmät ja toimittaa hermoja myös muihin aistinelimiin, jotka sijaitsevat pään, kirveen ja clavan välissä. Se on myös epätavallisesti kytketty ensimmäisen runko -segmentin ganglioihin . Kuitenkin, kuten niihin liittyvät niveljalkaiset, nämä ovat ensisijaisesti yhteydessä maanalaiseen ganglioniin leveillä hermosolmilla.
Tardigrade -aivot syntyvät useiden alun perin itsenäisten ganglionien fuusioinnista, jota voidaan tulkita viittaukseksi pään alkuperään vähintään kolmen segmentin fuusioinnista. Läheisesti sukua olevien niveljalkaisten aivot koostuvat myös useista ganglioneista, jotka muodostavat kolme aivojen aluetta, proto-, deuto- ja tritocerebrum. Siksi joskus oletetaan, että vastaavat rakenteet ovat homologioita , eli kudoksia, jotka palaavat tardigradin ja niveljalkaisten yhteisiin esi -isiin. Uudemmat elektronimikroskooppiset tutkimukset ovat ristiriidassa tämän näkemyksen kanssa ja päätyvät sen sijaan siihen, että tardigraden koko aivot vastaavat evoluutiolla niveljalkaisten protocerebrumia.
Kaksi vatsaan kulkevaa hermolankaa muodostavat rungon hermoston. Ne syntyvät maanalaisessa ganglionissa ja kulkevat rinnakkain eläimen takaosan kanssa. Jokaisessa hermojohdossa on neljä tai viisi gangliaa, jotka vastaavat neljää jalkaa kantavaa segmenttiä ja ehkä toinen jalkaton sukupuolielinten segmentti. Jalan segmentin kaksi ganglionia yhdistävät poikittaiset hermoyhteydet.
Tuntoelimet
Monilla, mutta ei kaikilla, tardigradesilla on punaiset silmät, joko punaiset tai mustat. Ne on suunniteltu niin kutsutuiksi pigmenttikuppioocelleiksi , mikä tarkoittaa, että kukin silmä koostuu kupin muotoisesta pigmenttisolusta, jota ympäröivät kaksi fotoreceptorisolua, joiden kovera puoli osoittaa kehon pintaa ja siten valoa. Ne saavat hermoja aivojen ulommat, takakehot, ja niiden erityisessä muodossa niitä voi esiintyä vain tardigradessa.
Lisäksi joillakin lajeilla on harjamaiset aistit rungon osissa , jotka oletettavasti reagoivat kemiallisiin tai kosketusärsykkeisiin. Langanmuotoiset aistit, cirri , löytyvät pään takaa, etenkin monissa merilajeissa , ja ne ovat luultavasti kosketuselimiä, kun taas clavae , hieman paksummat ja sisäisesti ontot langat, ovat todennäköisesti kemoreceptoreita . Monilla lajeilla on syylänmuotoisia kohoumia suun aukkojen, papillien , ympärillä , joilla todennäköisesti on myös tehtävänsä ympäristön havaitsemisessa.
Lisääntymiselimet
Tardigradessa on pohjimmiltaan vain yksi pariton sukurauha (gonadi), jota ympäröi todellinen kelomikudos, joka sijaitsee ruoansulatuskanavan yläpuolella ja kiinnittyy nivelsiteillä rungon takaosan seinämän etupäähän.
Flagelloidut siittiöt muodostuvat urosten kiveksiin . Siitä lähtee kaksi siittiöjohtoa, jotka yhdistyvät vatsaan seksuaalisen aukon kohdalla, gonopore, joka sijaitsee peräaukon edessä kehon keskiviivalla, usein putkimainen ulkoneva ja avoin ulkomaailmaan. Siemenportaiden toiminnallisesti selittämätön päällekkäisyys nähdään osoituksena sukurakon evoluutiosta.
Toisaalta naarasmunasarjassa on vain yksi munanjohdin, joka on lajista riippuen joko suolen oikealla tai vasemmalla puolella. Yhden luokan lajeilla, heterotardigradalla, sen suu sijaitsee erillisessä gonoporessa, joka yleensä sijaitsee peräaukon edessä, kun taas toisen luokan, eutardigradan, laji on takaosassa, josta tulee siten kloaakki . Monilla naarailla on yksi tai kaksi siemenrakkulaa, jotka parittelun aikana ottavat urosten siittiöitä ja varastoivat niitä munimiseen asti.
Hermafrodiittisilla yksilöillä on sukurauha, joka tunnetaan nimellä ovotestis , jossa sekä siittiöt että munasolut kypsyvät ja jotka voidaan vapauttaa jaetun munan tai siemenputken kautta.
jakelu
Tardigrades elää kaikilla mantereilla ympäri maailmaa, myös Etelämantereella, ja kaikissa valtamerissä. Niitä löytyy Keski -Euroopan sadevesikourista sekä säännöllisesti jäätyneiltä arktisilta altailta tai trooppisista sademetsistä , yli 6000 metrin korkeudessa Himalajalla , syrjäisillä saarilla, kuten Etelä -Sandwich -saarilla , 4690 metriä syvällä syvyydellä Intian valtameren pohjalla tai Atlantin keskellä ajelehtivilla ruskeilla levillä . Vaikka niitä esiintyy kaikilla ilmastovyöhykkeillä, pääasiallinen levinneisyysalue on napa- ja leutoilla leveysasteilla.
Luonnonmaantieteellisiä rakenne voi ainoastaan voidaan hyväksyä perheen ja suvun tasolla, joka voidaan liittää kanssa erottaminen vanhan mantereen Pangea osaksi Gondwana ja Lauraasia . Yhteensä kymmenen sukua ja 22 lajia ovat itse kosmopoliittisia eli niitä löytyy kaikkialta maailmasta. Niitä pidetään jäänteitä Pangea eläimistön, joka oli olemassa ennen geologinen epookki Triaskausi . Useimmilla muilla lajeilla on alueellisesti rajoitettu alue.
elinympäristö

Elinympäristönsä mukaan tardigrade voidaan pohjimmiltaan kuvata meren ( meren ), makean veden ( limninen ) tai maanpäälliseksi ( maanpäällinen ), jolloin kahden viimeisen luokan välinen ero on vain epäselvä, minkä vuoksi kollektiivista termiä limnoterrestrial käytetään usein . Kaikki tardigrades, vaikka jotkut ovat erittäin vastustuskykyisiä dehydraatiolle, ovat riippuvaisia ohuesta vesikalvosta aktiivisen elämän ajan.
Meren elinympäristö
Merilajit elävät sekä suolaisessa että murtovedessä, ja niitä voi esiintyä vuoroveden vyöhykkeeltä matalan vesialueen läpi valtamerien syville tasangoille; ainakin yksi laji kestää Marianan kaivanteen lattialla vallitsevan vedenpaineen . Jos merenpohja on mutainen, siellä olevat pohjaeläimet ovat enimmäkseen sopeutuneet ympäristöönsä vahvalla matonmuotoisella rungolla ja lyhentyneillä raajoilla; Toisaalta hiekkaisella tai kivisellä maaperällä, jossa on rakoja ja halkeamia, löytyy todennäköisemmin eläimiä, joilla on hyvin kehittyneet jalat. Yksi laji on ilmeisesti erikoistunut mangaani kyhmyt kuin elinympäristö, kun taas toiset laaja levät mattoja tai eläinten, kuten kivikoralleja (SCLERACTINIA), sammaleläinten , simpukat (Bivalvia), maasiirat (Isopodaa), siimajalkaisten (Cirripedia), merisiilit (Echinoidea) tai Sea nokat (Holothuroidea) elävät, osittain kommensaalin eli vaikuttamatta isäntä, mutta osin myös loistaudit . Ainakin yksi laji elää kelluvilla Sargassum -levillä Sargasso -meren avomeren keskellä . Maata kohti on selvä hiekkaraon eläimistö maaperän korkeimmilla senttimetreillä lähes kaikilla rannoilla, joilla eläimet elävät yksittäisten hiekkajyvien välissä. Jos lajeja on useita, ne jakautuvat useimmiten eri mikroympäristöihin, jotka voidaan erottaa toisistaan kosteus- tai lämpötilaeroilla. Siirtyminen maanpäällisiin muotoihin muodostuu niistä tardigradeista, jotka elävät meren jäkälissä , jotka ovat asettuneet lohkareisiin veden korkeuden yläpuolella, joka yleensä saavutetaan korkealla vuorovedellä ja joihin yleensä pääsee vain suolainen suihke.
Makean veden elinympäristö

Limniset lajit elävät sanan todellisessa merkityksessä virtaavissa vesissä sekä järvissä, lampissa, lampissa tai jopa yksittäisissä lätäköissä. Tardigrades ovat tavallisesti osa järven pohjassa elävää hiekkalaatikkoa; Löydöt jopa 150 metrin syvyydestä tunnetaan. Eläimet elävät enimmäkseen maaperän ylimmällä, happipitoisella senttimetrillä, järven rannalle, missä löysä hiekka voi vahingoittaa eläimiä vakavasti, mutta enimmäkseen hieman syvemmälle. Levät ja vesikasvit ovat myös kolonisoituja, ja tardigradesia voi toisinaan löytää vesililjoista järven pinnalla .
Usein yksittäiset yksilöt voidaan jäljittää sadevesikouruihin; eläimet pestään oletettavasti sammalilla katolla, joten niitä pidetään maanpäällisinä. Kuumat lähteet muodostavat myös makean veden elinympäristön, johon jotkut lajit ovat asuneet.
Elinympäristö

Tärkeimmät maanpäälliset elinympäristöt ovat sammal nurmikot; siellä elävät lajit tunnetaan sammalta rakastavina tai bryofiilisinä . Koska sammalit varastoivat vettä välivälille, niitä voidaan kuitenkin pitää myös vesieliöintinä. Usein sammaleiden eri vyöhykkeillä on erilaisia lajeja; risoidi-kerroksen tardigrade-eläimistö, jonka kanssa sammalat ovat juurtuneet maaperään, ja esimerkiksi fotosynteettisesti aktiivinen, mutta kuivumisaltis ulkokerros eivät ole identtisiä. Lisäksi eläimet löytyvät jäkälien tai sopivassa kukkivat kasvit kuten bromeliads (Bromeliaceae), syöttäjä kasvit (kannukasvit) in saxifrage ( Saxifraga ) Mannsschild ( Androsace ) tai karstaamalla ( Dipsacus ). Kaikissa näissä kasveissa vesi kerääntyy lehtien juuriin tai muihin kupin muotoisiin kasvinosiin; niistä tulee pienikokoinen elinympäristö.
Tardigradesia esiintyy usein myös metsien lehtihiekassa tai itse maassa, ja pyökkimetsät näyttävät olevan erityisen suosittuja. Hieman epätavallisempi elinympäristö ovat korkeiden vuoristojen jäätiköt : siellä tummat pölykerrostumat tai hienorakeiset roskat voivat johtaa lämmön imeytymiseen päivän aikana ja siten pinnan väliaikaiseen sulamiseen; tardigrade löytyy myös tuloksena olevasta vesipitoisesta kryokoniittikerroksesta . Kaupunkien elinympäristöjä ei ole juurikaan tutkittu tähän mennessä.
Tardigrades ovat aktiivisia vain, kun ne on peitetty ohuella vesikalvolla substraatissaan, joten ne eivät voi elää kuivilla alueilla, kuten aavikoilla . Monet maanpäälliset lajit ovat kuitenkin erittäin kuivuutta kestäviä ja voivat siksi selviytyä säännöllisistä kuivista kausista ja äärilämpötiloista. Tämä perustavanlaatuinen ekologinen etu, jonka he jakavat joidenkin rotifareiden (Rotifera) kanssa, voivat parhaiten pelata kilpailevia eläimiä, kuten sukkulamatoja (Nematoda) vastaan, joissa ympäristön kosteuspitoisuus vaihtelee voimakkaasti, mikä on ainakin osittain etusijalla monista tardigrade -lajeista Sammal ja jäkälät selittivät, jotka kuivuvat hyvin lyhyessä ajassa tai voivat tulla veden päälle. Näiden elinympäristöjen asuttaminen, joihin harvat muut eläimet pääsevät, on tärkeä syy tardigradin suurelle evoluutiotehtävälle.
Väestötiheydet
Tarkkoja tietoja populaatiotiheydestä on saatavana vain muutamille lajeille ja toistaiseksi vain näytteinä: Hiekkarannoilla oli jopa 35 yksilöä kuutiosenttimetriä hiekkaa kohti, maaperässä laskettiin jopa 30 yksilöä neliösenttimetriä kohti, kun taas sammalta tyynyt, joissa on noin 200 yksilöä per odotukset, neliösenttimetrit edustavat erittäin tiheästi asuttua elinympäristöä. Yleensä väestötiheys on kuitenkin selvästi tämän enimmäisarvon alapuolella. Siltä osin kuin voidaan sanoa toistaiseksi riittämättömän tietotilanteen perusteella, merilajien yksilölliset määrät ovat yleensä paljon pienempiä kuin maa- tai makean veden muodot.
Väestötiheyteen voivat vaikuttaa lämpötila ja kosteus, ravinnon tarjonta, loisten ja saalistajien lukumäärä tai ympäristön epäpuhtaudet, ja maanpäällisissä lajeissa ne vaihtelevat enimmäkseen vuodenajan mukaan, ja ensimmäinen huippu on keväällä ja alkukesällä ja toinen huippu syksyllä.
Kuljetus ja liikkuminen
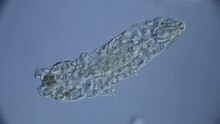
Tardigrade leviää vain hyvin vähäisessä määrin aktiivisen liikkumisen kautta; useimmat eläimet tuodaan sen sijaan uusiin mahdollisiin elinympäristöihin tuulen, veden tai eläinten toimesta. Tämä passiivinen kuljetus vaikuttaa ensisijaisesti eläinten muniin sekä kystat ja tynnyrit - elämänvaiheet, jotka ovat suurelta tai äärimmäiseltä määrin riippumattomia ympäristön ääripäistä.
Merilajeja voidaan ajautua valtameren virtauksiin, mikä todennäköisesti auttaa heitä erityisillä purjeen kaltaisilla pullistumilla niiden ulkokuorella. Maan ulkopuolisia lajeja kuljetetaan toisinaan samalla tavalla ylivuotavien purojen tai sulamisveden kautta. Väliaikaiset kuivat elinympäristöt mahdollistavat munien tai kystien leviämisen tuulen mukana ( anemokorinen ), kun taas pienempiä osuuksia voidaan silittää vesiroiskeissa. Grönlannin myrskyn aikana jopa aikuisia eläimiä havaittiin putoavista sadepisaroista, jotka myrsky todennäköisesti heitti ylös etukäteen. Oletettavasti hyönteiset tai linnut, joihin munat tai kystat tarttuvat, tuovat myös tardigradesia uusiin elinympäristöihin ( eläintarhoihin ).
Aktiiviseen liikkumiseen kaikki lajit tarvitsevat ohuen ympäröivän vesikalvon. Sitten he käyttävät kolmen ensimmäisen runkosegmentin jalkoja, joita voidaan toisin kuin vastaavaa kolumbusta liikuttaa paitsi pareittain, myös yksitellen, esimerkiksi ryöstää hiekkajyvien yli tai kiivetä sammal tyynyissä. Jalkojen kynsiä tai liimakiekkoja käytetään tarttumaan vastaavaan alustaan. Toisin kuin kolme ensimmäistä jalkaparia, kaksi takaraajaa tukeutuvat maahan ja joissakin lajeissa myös taaksepäin; ne on mukautettu tähän muutettuun toimintoon eri kynsien kohdistuksen avulla. Suurin kokeellisesti mitattu juoksunopeus on 17,7 senttimetriä tunnissa; arvo tekee ryhmän latinalaisesta nimestä Tardigrada , joka on johdettu latinalaisesta tarduksesta 'hidas' ja gradus 'askel', joka tarkoittaa 'hidasta juoksijaa'.
Valotaksista eli liikettä valonlähteisiin ja niistä pois ei ole vielä tutkittu riittävästi. Nuoret eläimet näyttävät reagoivan negatiivisesti fotokineettisesti , toisin sanoen ne reagoivat valosäteilyyn nopeammilla liikkeillä ja spontaanilla suunnanmuutoksella ilman erityistä välttelyreaktiota. Koska valonsäteily liittyy usein lämpösäteilyyn ja sen jälkeiseen veden haihtumiseen, tämä käyttäytyminen liittyy ehkä kuivumisriskiin, joka on uhkaavampi nuorille eläimille.
Merilaji pystyy uimaan aktiivisesti; sen kynsinauha on kellon muotoinen ja voi karkottaa suunnatun vesisuihkun supistumalla; eläimet liikkuvat kuin meduusat takaiskuperiaatteen mukaisesti.
ravintoa

Useimmat tardigrades ruokkivat kasvisruokavaliota, pääasiassa leväsoluja , joita he löytävät joko luonnosta tai jäkälistä. Sammalien kasvisolut sitä vastoin kuuluvat harvoin ravintoon, jopa siellä pysyvästi elävissä eläimissä. Levien lisäksi maaperässä elävät lajit nielevät myös orgaanista jätettä yhdessä sen sisältämien bakteerien ja sieni-itiöiden kanssa ; jotkut muodot elävät kuitenkin myös kokonaan tai osittain saalistavasti. Niiden valikoima saalis sisältää alkueläinten , rataseläimet (Rataseläimiä) ja sukkula- (Nematoda), mutta myös muita karhukaiset. Jotkut merilajit elävät ektoparasiiteina saksanpähkinöiden tai korvien iholla . Maalla tardigrades on sopeutunut huonosti loiseen elämäntapaan jatkuvan kuivumisen riskin vuoksi; täällä tunnetaan vain yksi laji, joka voi olla endoparasiittinen maa -etanoissa .
Syödäkseen tardigrade painaa suunsa kartiota vasten vaurioitunutta kasvisolua, saaliin ihoa tai isännän kehon seinämää. Edistämällä neulateräviä tyylejä ne lävistetään tai porataan ja koko solu tai kehon sisältö imetään pois. Erityisen suuret lajit eivät sitä vastoin voi ottaa vain nestemäistä ruokaa, vaan myös imeä saaliinsa kokonaisuutena; tämä vaikuttaa erityisesti pienempiin pyöräeläimiin ja tardigradiin.
Petoeläimet, loiset ja kommensaalit
Karhukaiset tärkeimmät saalistajat ovat muita karhukaiset lajeja, rataseläimet (Rotatoria) ja ankeroiset, sekä punkkeja (Acari), hämähäkit (Araneae), hyppyhäntäiset (Collembola), eri hyönteisten toukkia , mutta myös epäspesifisesti "laiduntaminen" organismeja, kuten lieroille (lierot) tai erilaisia makean veden rapuja.
Jotkut tardigradit joutuvat lihansyöjien sienien uhreiksi, joiden hienot solukierteet ( hyphae ) ovat kietoutuneet silmukoihin, joihin heidän saaliinsa tarttuvat, ja jotka sitten kasvavat pyydystetyiksi eläimiksi. Muista sienistä, kuten Harposporium , suvuton vaiheessa rihmasienen suvun Atricordyceps antavat konidioita, suvuton itiöitä, jotka todennäköisesti kun ne syövät karhukaiset, itää suolistossa ja sulattaa niiden saaliin sisällä. Sieni voi kuitenkin tunkeutua eläinten kynsinauhaan myös ulkopuolelta. Muita tärkeitä tardigraden sieni -loisia ovat potta -sieni Sorochytrium milnesiophthora ja ikeen sienet Ballocephala sphaerospora ja Ballocephala verrucospora .
Oletettavasti alkueläinten joukossa on useita loisia ; kunnes se on tutkittu yksityiskohtaisesti vielä, mutta ripsieläimiä Pyxidium tardigradum että edullisesti tartuttaa karhukaiset. Se elää oletettavasti symphoriontina , mikä tarkoittaa, että vain isäntä voi levittää sitä, mutta se ei näytä olevan vaarallinen itselleen.
Monia symbioottisia bakteereja löytyy usein meren tardigradista ; Maalla elävillä muodoilla on myös enimmäkseen runsas bakteeriperäinen suolistofloora. Proteobacterium Xanthomonas campestris , tärkeä tuholaisten kasveja, uskotaan lähetetään karhukaiset.
Molting
Tardigrade voi kasvaa kooltaan vain säännöllisellä sulatuksella . Ei-soluinen ulompi iho (kutikulaarinen) yhdessä stilettojen, etu- ja takarakon vuorauksen ja jalkojen kynnet irtoavat. Hieronta alkaa aina eläinten etupäästä, sillä vanhat tikat ja suuontelon ja esilihan kynnet karkotetaan. Kun ruumis supistuu tilapäisesti, se irrottaa itsensä vanhasta ihosta, joka sitten riisutaan ja jätetään tyhjäksi irtoavaksi paitaksi ( exuvium ). Tämän prosessin aikana, joka kestää tavallisesti noin 5–10 päivää, eläimet ovat yksipuolisessa vaiheessa , ilman niittejä , jolloin ne eivät voi syödä ruokaa. Sulkemisen aikana alla olevasta orvaskedestä muodostuu uusia kynsinauhoja, kun taas uudet tikat syntetisoidaan sylkirauhasissa ja kynnet rakennetaan erityisillä kynsirauhasilla.
Yksittäisillä yksilöillä voidaan havaita jopa 13 nuhaa elämänsä aikana; Sen lisäksi, että ne mahdollistavat elinikäisen kasvun, ne vähentävät myös kehon sisäistä painetta, hylkivät kynsinauhoissa olevia jätemateriaaleja tai poistavat loisia. Monet lajit munivat munansa huopapaitoihin.
Vastusvaiheet
Monet tardigradit ovat kehittäneet ainutlaatuisia mukautuksia selviytyäkseen kuivuudesta, kylmyydestä, veden suolapitoisuuden jyrkistä vaihteluista tai hapen puutteesta. Panostus kestävyyttä on niiden Eutelia: Kun alkion kehitys, solunjakautumisen ja sen vaativissa vaiheissa, tuskin tapahtuu (paitsi iturataan ). Tämä johtaa jo erittäin korkeaan säteilynkestävyyteen (lukuun ottamatta steriiliyttä ), vertailua varten katso taulukko kohdassa → Säteilyvauriot . Lisäksi jotkut lajit sopeutuvat vuodenaikoihin morfologisten muutosten kautta, kun taas toiset voivat muodostaa paksuseinäisiä kystat. Äärimmäinen sopeutumismuoto on kuitenkin niin kutsuttu kryptobioosi , jossa eläimet menevät kuolemaan lähellä olevaan tilaan, jossa ei voida enää rekisteröidä aineenvaihduntaa. Kaikki resistenssivaiheet auttavat selviytymään epäsuotuisista ympäristöolosuhteista ja antavat siten tardigradille kehittyvän kilpailuetun muihin eläinryhmiin nähden. Niillä on myös rooli eläinten passiivisessa kuljetuksessa uusiin mahdollisiin elinympäristöihin. Tardigrade on myös ensimmäinen eläin, jonka tiedetään pystyvän selviytymään avaruudessa.
Syklomorfoosi
Syklomorfoosi on kehon muodon säännöllinen muutos vastauksena suhdannevaihteluihin ympäristöolosuhteissa. Suurin osa eläinten elämänvaiheista, joita kutsutaan morfeiksi , vaihtuvat vuodenajasta riippuen. Toistaiseksi cyclomorphosis on vain tunnettu meren karhukaiset suvuista Halobiotus , Amphibolus ja Hypsibius . Tähän mennessä parhaiten tutkittu esimerkki löytyy lajista Halobiotus crispae , joka elää Grönlannin vuorovesialueella : Täällä pseudosimplex -niminen talvimorfia voidaan erottaa kesämorfista . Edellinen kykenee kestämään pakkasen lähellä olevia lämpötiloja ja pysyy liikkuvana myös alhaisissa lämpötiloissa, mutta se on steriili; vain paljon vähemmän vastustuskykyinen kesämorfia pystyy lisääntymään.
Kystat
Erityisesti makeassa vedessä elävät lajit, mutta myös sammalten tai lehtien kuivikkeet, voivat kehittää vastustuskykyä, joka tunnetaan kystinä. Tätä varten eläimet supistuvat 20-50 prosenttiin aiemmasta kehon koostaan, vähentävät aineenvaihduntaa ja joissakin tapauksissa myös hajoavat sisäelimiä. Tähän prosessiin liittyy enintään kolme epätäydellistä, välittömästi peräkkäistä hierontaa, joiden lopussa eläintä ympäröi kynsinauhakerroksista tehty moniseinäinen päällyste. Tässä tilassa eläimet voivat selviytyä yli vuoden. Heti kun ympäristöolosuhteet ovat muuttuneet, eläimet voivat vapautua kotelostaan jälleen 6–48 tunnin kuluessa.
Pohjimmiltaan kystat muodostuvat vain vedessä; ne eivät ole yhtä kestäviä kuin jäljempänä mainitut rummut, ja toisin kuin ne, vesipitoisuutensa vuoksi ne eivät myöskään ole lämmönkestäviä.
Anoksibioosi
Anoksibioosi on termi, jota käytetään kuvaamaan hapen puutteen sietokykyä. Jos tämän hengityskaasun pitoisuus on liian pieni, osmoregulaatio hajoaa , eläimet nauttivat vettä ja turpoavat merkittävästi. Koko aineenvaihdunnan on nyt tapahduttava anaerobisesti, myrkylliset jätteet kerääntyvät kehon onteloon. Useimmat tardigrades pystyvät edelleen selviytymään tällaisesta tilasta 3-5 päivää; jopa kerrotaan, että yksittäiset Echiniscoides -suvun yksilöt kykenivät selviytymään jopa kuusi kuukautta suljetussa koeputkessa, jossa oli mädäntynyttä. Paluu normaalitilaan kestää muutamasta minuutista muutamaan tuntiin anoksibioottisen vaiheen kestosta riippuen.
Anoksibioosi on erityisen tärkeä elinympäristöissä, joissa happipitoisuus voi laskea tilapäisesti jyrkästi, esimerkiksi suurissa staattisissa vesimassoissa. Tardigrade, joka elää levien matoilla vuoroveden vyöhykkeellä, altistuu myös äärimmäiselle hapenpuutteelle laskuveden aikaan, ja evoluution kannalta he olivat ehkä ensimmäisten joukossa.
Osmobioosi
Osmobioosi on kyky sietää veden suolapitoisuuden vaihteluita. Tardigrade, kuten useimmat eläimet, ovat elinkelpoisia vain, jos ionien pitoisuus vedessä on tietyissä rajoissa. Erityisesti vuorovesialueella elävät lajit voivat kuitenkin selviytyä hämmästyttävän suurista suolapitoisuuden vaihteluista lyhyen ajan. Monet lajit muodostavat alla kuvatut pienet tynnyrit hyvin suolaisessa vedessä; Tämä on kuitenkin erityistapaus alla selostetulle anhydrobioosille ja suora vastaus sisäiseen kuivumiseen, joka liittyy ympäristön korkeaan suolapitoisuuteen. Tynnyrin muodostumista ei näin ollen pidetä muodollisesti osmobioottisena ilmiönä.
Cryobiosis
Cryobiosis on kyky kestää matalia lämpötiloja. Se on kryptobioosin ilmentymä, jolle on ominaista melkein täydellinen aineenvaihdunnan pysähtyminen. Tapahtuu niin sanottu lepotila . Alla tarkemmin kuvattu tynnyrien muodostus on mahdollista, mutta ei pakollista.
Kun lämpötila laskee hitaasti, aineenvaihdunta muuttuu vähitellen, koska useimmat entsyymit menettävät aktiivisuutensa ja sen sijaan vaikuttavat matalan lämpötilan aktiiviset katalyytit, jotka syntetisoivat sokeritrehaloosia ja muita kylmäsuojaavia aineita: Nämä suojaavat herkkää biokalvoa ja korvaavat olennaiset molekyylit sitoutuneen veden . Erityisesti solunulkoisessa kehon nesteessä aiemmin tunnistamattomat, keskipainoiset molekyylit, jotka toimivat jäädyttävinä bakteereina, stimuloivat jääkiteiden hallittua kasvua, jotka jäätymisenestoproteiinit stabiloivat pieninä.
Tällä tavalla eläimet voivat selviytyä pakkasen alapuolella olevista lämpötiloista. Tämä kyky mahdollistaa vain napa -alueiden ja korkeiden vuorijäätiköiden asuttamisen, joissa ympäristön lämpötila laskee säännöllisesti jäätymispisteen alapuolelle. Esimerkiksi alalaji Echiniscus sigismundi groenlandicus selviää talvijäässä jäädytettynä 6–8 kuukautta ja sietää säännöllistä sulamista ja jäätymistä vuorovesistä siirtyessä .
Tardigrades selviytyi jopa kymmenen päivää vapaassa tilassa. FOTON-M3- satelliitin avulla ESA paljasti useita avaruudessa esiintyviä tardigrade-näytteitä tyhjiölle, kylmälle ja UV-säteilylle vuonna 2007. Palattuaan tiedemiehet löysivät selviytyjiä jopa äärimmäisissä olosuhteissa altistuneista tardigradeista.
Koska tardigrades voi selviytyä äärimmäisistä - 273 celsiusasteen lämpötiloista, jos ne jäähtyvät riittävän hitaasti, on epäilty niiden olevan maan ulkopuolisia elämänmuotoja . Tällaisia lämpötiloja ei koskaan edes saavutettu luonnollisessa elinympäristössään, eikä kyky näin olisi voinut syntyä maan päällä luonnollisen valinnan kautta . Tätä näkemystä vastustaa se tosiasia, että elävä olento, joka selviytyy maan pakkaslämpötilojen alapuolella korvaamalla kudoksensa vesi suoja -aineilla, kuten trehaloosilla, ja kontrolloimalla itse jäätymisprosessia, voi mahdollisesti selviytyä -273 celsiusasteen lämpötiloista ilman lisämuutoksia. Tardigrades ovat monisoluisia polyekstremofiileja .
Anhydrobioosi
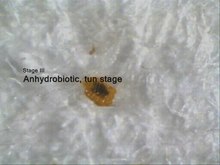
Anhydrobioosi Richtersius -koroniferin esimerkin avulla .
Vaihe 1: edelleen hydratoitu ja aktiivinen dehydratoituna.
Vaihe 2: 10 minuutin kuluttua jalat ovat kuivuneet ja vetäytyneet sisään.
Vaihe 3: tynnyrivaihe (anhydrobioosi).
Vaihe 4: palauttaminen veden lisäämisen jälkeen.
Anhydrobioosi on kyky selviytyä kehon kuivumisesta raskaan veden menetyksen kautta. Sitä esiintyy lähes kaikissa maalajeissa ja se liittyy lieriömäisten, liikkumattomien vastustusvaiheiden, tynnyrien, muodostumiseen . Koska aineenvaihduntaa ei voida havaita tynnyreissä anhydrobioottisessa tilassa, anhydrobioosi kuuluu myös yleiseen termiin kryptobioosi.
Joillakin lajeilla monet yksilöt kokoontuvat ennen varsinaista tynnyrinmuodostusta ja muodostavat löysän pallon. Tätä aggregaatiota pidetään käyttäytymissopeutumisena ja se todennäköisesti suojaa sisällä olevia eläimiä hieman enemmän ympäristövaikutuksilta, joten kuivumisprosessi on heille hieman hitaampi - liian nopea kuivuminen voi myös johtaa kuolemaan tardigradessa.
Tynnyrin muodostuminen alkaa morfologisista reaktioista: jalat vedetään sisään ja kehon koko pinta vähenee huomattavasti. Lipidejä , rasvaliukoisia aineita, joiden on muun muassa tarkoitus suojata sieni-iskuilta, vapautuvat yhä enemmän kynsinauhojen huokosten kautta . Seurauksena on vaiheen muutos näiden lipidien, kynsivalliin yhtäkkiä tulee vettä läpäisemättömiä, jonka tiettynä ajankohtana. Tämä vähentää veden haihtumista ja mahdollistaa valmistusajan pidentämisen jälleen anhydrobioottiseen tilaan, jonka aikana suojayhdisteitä on syntetoitava.
Suurin ongelma anhydrobioosissa on se, että tärkeiden makromolekyylien, kuten proteiinien , fosfolipidien tai nukleiinihappojen, rakenteellinen eheys on säilytettävä hinnalla millä hyvänsä, muuten solut joutuisivat peruuttamattomiin vaurioihin. Suurinta osaa näistä yhteyksistä ympäröivät löyhästi kiinnittyneet vesimolekyylit, joiden menetys aiheuttaisi hallitsemattomia reaktioita niiden välillä. Ratkaisu ongelmaan, jonka tardigrade on löytänyt evoluutiohistoriansa aikana, on korvata sidottu vesi dehydraation aikana muilla yhdisteillä, jotka voidaan helposti hajottaa uudelleen nesteytyksen aikana (veden kostutus). Tärkein näistä yhdisteistä on sokeri trehaloosi , jota tuotetaan suurina määrinä aikana valmisteltaessa anhydrobiotic tila ja joskus väkeviä kertoimella 23 verrattuna alkuperäiseen tilaan. Se ei ainoastaan suojaa biokalvoa ja estää reaktioita dehydratoituneiden proteiinien ja muiden solukomponenttien, kuten hiilihydraattien välillä, vaan estää myös hallitsemattoman hapettumisen , mikä voi myös tuhota tärkeitä makromolekyylejä. Uudet tutkimukset viittaavat siihen, että tardigradesilla on uudentyyppinen häiriintynyt proteiini , joka kuivutettuna muodostuu ja lasittuu yhä enemmän , eli muodostaa lasimaisen suojamateriaalin. Tämän proteiiniperheen ja trehaloosin lisäksi glyseriinillä on todennäköisesti myös rooli näissä prosesseissa. Samaan aikaan lipidipisaroiden tehtävänä on pitää kehon kudokset erillään toisistaan, jotka normaalitilassa eivät ole vaarassa päästä liian lähelle, mutta tynnyritilassa joutuvat kosketuksiin ja voivat aiheuttaa ei -toivottuja silloitusreaktioita. Kestää noin 5-7 tuntia, ennen kuin eläimet palaavat normaalitilasta vakaaan tynnyrivaiheeseen.
Heti kun eläimet ovat saavuttaneet anhydrobioottisen tilan, aineenvaihdunnan aktiivisuutta ei enää voida havaita, ainakin kuivassa ympäristössä; erityisesti hapen kulutus laskee nollaan. Tässä vaiheessa eläimet voivat selviytyä äärimmäisestä kuivumisesta, upottamisesta suolaliuokseen, eetteriin , puhtaaseen etanoliin tai nestemäiseen heliumiin , lämpötiloihin -196 ° C - +90 ° C, korkeaan hydrostaattiseen paineeseen ja tyhjiöön .
Heti kun vettä on jälleen tarpeeksi, muutos takaisin normaalitilaan, palautus tapahtuu. Tätä varten trehaloosi hajotetaan aerobisesti eli käyttämällä ilmakehän happea ja korvataan sitten vedellä. Palautumisaika riippuu ympäristön lämpötilasta, happipitoisuudesta ja pH -arvosta sekä kryptobioottisessa tilassa vietetystä ajasta ja vaihtelee jonkin verran myös lajeittain. Useimmissa tapauksissa se on kymmenestä minuutista useaan tuntiin, ja jos tynnyri on erittäin pitkä, jopa muutama päivä.
Se, onko paluu aktiiviseen elämänvaiheeseen onnistunut, riippuu ensisijaisesti tynnyrinmuodostuksen oikeasta järjestyksestä ja eläinten ravitsemustilasta. Erityisen tärkeitä tässä yhteydessä ovat lipidivarat, joilla ei ole pelkästään roolia kryptobioottisessa tilassa, kuten edellä mainittiin, mutta ne voidaan myös muuntaa glyseroliksi ja trehaloosiksi ja tarjota myös energiaa palautusprosessille. Niin kauan kuin eläimillä on mahdollisuus täydentää näitä varastoja säännöllisin väliajoin ja lisäksi kaikki kuivuminen tapahtuu vain hitaasti, ne voivat vaihtaa aktiivisesta tilaan anhydrobioottiseen tilaan ja päinvastoin säännöllisin väliajoin. Esimerkiksi vuorovesivyöhykkeellä elävät Echiniscoides- suvun yksilöt voidaan kokeellisesti asettaa kuuden tunnin jaksoon.
Tardigrade kryptobiotisessa tilassa herättää kysymyksiä elämän määritelmästä : aineenvaihdunta, kasvu ja lisääntyminen mainitaan usein elävän organismin ominaispiirteinä. Mitään näistä ominaisuuksista ei kuitenkaan löydy tynnyrivaiheessa, joten paluuta aktiiviseen tilaan voitaisiin kuvata "ylösnousemukseksi kuolleista" 1700 -luvun italialaisen luonnontieteilijän Lazzaro Spallanzanin sanoin. Toisaalta, vaikka monet, mutta eivät kaikki, tardigrade palaavat anhydrobioottisesta vaiheesta aktiiviseen tilaan, tardigradologi John H.Crowe kysyi vuonna 1975:
- Tarkoittaako tämä sitten, että he "kuolivat" ollessaan "kuolleita"?
- Tarkoittaako se sitten sitä, että he "kuolivat" ollessaan "kuolleita"?
Crowen mielestä elämän määritelmää ei voida rajoittaa yksittäisiin piirteisiin; Sen sijaan hän näkee elävän järjestelmän rakenteellisen jatkuvuuden ja eheyden jatkuvan olemassaolon "elintärkeänä" sanan kirjaimellisessa merkityksessä.
Lisääntyminen ja elinkaari
Tardigrade voi lisääntyä sekä aseksuaalisesti että seksuaalisesti. Useimmat lajit lisääntyvät yksinomaan seksuaalisesti.
Suvuton lisääntyminen
Aseksuaalinen lisääntyminen on mahdollista vain partenogeneesin eli hedelmöittymättömien munien kypsymisen jälkeen, jotka myöhemmin kehittyvät naaraiksi. Sitä esiintyy useissa lajeissa, mukaan lukien Echiniscus- ja Pseudoechiniscus -suvut . Uroseläimiä ei usein tunneta, vaikka kääpiö -uroksia voitaisiin joissakin tapauksissa kuvata jälkikäteen. Näillä ja muilla lajeilla voidaan määrittää partenogeneettisen ja seksuaalisen lisääntymisen rinnakkaiselo; urokset kykenevät periaatteessa erottamaan sopivat, seksuaalisesti aktiiviset, sopimattomat, partenogeneettisesti lisääntyvät naaraat. Partenogeneesiin liittyvä tärkeä etu on ponnistelujen puute kumppanin löytämisessä, vakava haitta on geneettisen monimuotoisuuden väheneminen. Tämän kielteisiä seurauksia kuitenkin osittain kompensoi kyky yksinkertaisesti välttää haitallisia ympäristövaikutuksia kryptobioosin avulla.
Seksuaalinen lisääntyminen
Seksuaalinen lisääntyminen tardigradessa johtaa useimmiten, mutta ei aina, kahden sukupuolen olemassaoloon. Jotkut hermafrodiittiset (hermafrodiitti) lajit tunnetaan, jotka kykenevät hedelmöittymään itsestään; Muna- ja siittiösolut kypsyvät samassa sukurauhasessa. Niillä on myös se etu, että heidän ei tarvitse kuluttaa energiaa kumppanin etsimiseen, mutta ne eroavat partenogeneettisistä yksilöistä, jotka lopulta kloonaavat itsensä geneettisen rekombinaation mahdollisuuden kautta .
Suurimmalle osalle lajeista on kuitenkin eri sukupuolet, joita ei kuitenkaan aina ole helppo erottaa ulkopuolelta. Jos väestörakenteesta on tehty yksityiskohtaisia tutkimuksia, naisia on lähes aina selvästi enemmän.
Hedelmöitys voi tapahtua naaraspuolisen kehon ulkopuolella ja sisällä, naaraspuolisten munasolut liittyvät enimmäkseen molttiin. Molempien prosessien yksityiskohdat riippuvat muun muassa eläinten elinympäristöstä.
Merilajeissa urosten siittiöt yleensä laskeutuvat naaraiden siemennesteisiin, jotka sitten munivat munat vapaasti ja kiinnittyvät substraattiin, kuten leväsoluihin. Lisäksi sattuu myös, että naaras munii hedelmöittämättömät munansa heti heittämisen jälkeen hylättyyn kynsinauhaan, jossa uros hedelmöittää ne sitten ulkoisesti.
Maan ulkopuolisilla lajeilla siittiöitä ruiskutetaan toisinaan vanhan ja uuden ihon väliseen rakoon ennen naaraan sulamista tai sen aikana; heti kun naaras munii munansa hylättyyn paitahoitoon, hedelmöitys tapahtuu. Erityisesti maanpäällisten lajien kanssa tapahtuu usein parittelu, jonka vuoksi uros tarttuu naaraspuoliseen; Tällaista yhteyttä ei hylätä edes seksuaalisen toiminnan ulkoisten häiriöiden yhteydessä. Uros tuo nyt siittiöitä naisen sukupuolielimiin ennen naaraan lopettamista. Vaihtoehtoisesti siittiöitä voidaan myös sijoittaa siemennesteisiin tai injektoida suoraan ulkoisen ihon läpi kehon onteloon; jälkimmäisessä tapauksessa hedelmöitys tapahtuu naarasrauhasessa.
Munittujen munien määrä vaihtelee lajista riippuen 1 ja 35 välillä, ja tämä määrä kasvaa tasaisesti iän myötä. Yksinäiset naaraat voivat elinkaaren aikana tuottaa yli 100 munaa. Merilajeissa ne ovat ohuita tai paksuja kuoria ympäristöolosuhteista riippuen, kun taas maalajeilla voidaan havaita erilaisia munanmuotoja: Sileät munat kerrostuvat enimmäkseen naaraspuolisten paitapuseroihin, kun taas munat, joissa on paksuja, hienostuneesti koristeltuja ja oletettavasti kuivumista -kestävät kuoret ovat vapaita sammalkasveista tai kuoripaloja liimataan. Jotkut makean veden lajit käyttävät hyönteisten tai muiden niveljalkaisten irrotettuja eksoskelettoreita säilytykseen.
Erityisiä käyttäytymissopeutumisia lisääntymisen aikana havaitaan vain harvoissa tapauksissa: joissakin lajeissa on havaittu primitiivinen ”pariutumisrituaali”. Uros silittää naaraansa Cirri -pään päällä. Jonkin ajan kuluttua tällä tavalla stimuloituna naaras munii munansa hiekanjyvälle, jolle uros sitten kerää siittiöitä. Se, että jotkut naaraat vetävät irtoavia paitoja, joihin he ovat munineet, jonkin aikaa tulkitaan yksinkertaiseksi tapaukseksi "sikiönhoidosta".
Jos olosuhteet ovat suotuisat, poikaset kuoriutuvat noin 5–40 päivän kuluttua. Tätä varten ne imevät nestettä ympäristöstä, kunnes niiden laajeneva runko rikkoo munankuoren. Jotkut lajit rikkovat myös munankuorta suullaan. Koko prosessi kestää yleensä vain muutaman minuutin.
Nuoret eläimet ovat enimmäkseen värjäämättömiä ja niillä on vähemmän harjaksia, kirveitä tai kynsiä kuin aikuisilla. Muuten ne ovat hyvin samankaltaisia kuin nämä, joten niiden kehitys tapahtuu suoraan eli ilman toukkavaihetta. Joillakin Heterotardigrada -luokan lajeilla peräaukko ja sukuelinten aukko muodostuvat vasta yhden tai kahden hampaan jälkeen; Tätä tulkitaan joskus viittauksena epäsuoraan kehitykseen. Useimmissa tapauksissa nuoret eläimet kasvavat vain lisäämällä yksittäistä solutilavuutta, mutta eivät lisäämällä solujen määrää, niin että monissa kudoksissa on jo kuoriutumisen jälkeen aikuisen muodon lopullinen solumäärä. Mitooseja (solujakautumisia) voidaan kuitenkin joskus havaita myöhemmissä vaiheissa , mikä oletettavasti palvelee kuolleiden solujen korvaamista. Joka tapauksessa seksuaalinen kypsyys saavutetaan vasta useiden erien jälkeen.
Tardigraden normaali käyttöikä on kolme kuukautta ja kaksi ja puoli vuotta; se vastaa useimpien merilajien todellista elinikää. Maan ulkopuolisissa lajeissa eläinten elämä kuitenkin joskus tai usein keskeytyy kryptobioottitiloilla, joiden aikana sairaat yksilöt eivät vanhene. Tällä tavalla sammalta elävät lajit saavuttavat usein todellisen elinkaaren, joka on neljä vuotta tai enemmän; yksittäistapauksissa ne voivat myös kestää vuosikymmeniä. Kasvitieteellisen museon kuivuneesta sammalta löydettiin tardigrade, joka 120 vuoden kuluttua voitaisiin "elvyttää" anhydrobioottisessa tilassa.
Tardigrade ja mies
Tardigradea esiintyy lähes kaikissa ihmisten elinympäristöissä, mutta ne ovat tuskin havaittavissa pienen kokonsa ja epätavallisen elämäntapansa vuoksi. Koska niillä ei ole välitöntä taloudellista, lääketieteellistä tai eläinlääketieteellistä merkitystä, ne ovat useimmille ihmisille tuntemattomia. Ne, jotka näkevät ne ensimmäistä kertaa, kuvailevat niitä usein "söpöiksi", adjektiiviksi, joka löytyy hyvämaineisista eläintieteellisistä julkaisuista ja joka on luultavasti innoittanut paitsi karhun kaltainen ruumiin muoto, myös eläinten kömpelö liikkuminen . Niiden pitäminen yksitoikkoisessa viljelmässä eli yhdessä vähintään yhden muun lajin kanssa on mahdollista.
Vaaratietoja ei ole saatavilla; yksi laji, Thermozodium esakii , on kuitenkin saattanut kuolla sukupuuttoon. Koska tardigrade on erittäin herkkä ympäristömyrkkyille , erityisesti rikkitrioksidille , on ideoita käyttää eläimiä paikan ympäristön laadun indikaattoreina. Unkarilaiset tutkijat esittivät tutkimuksia Tardigradan vahingoittamisesta sammaleiden raskasmetallisaasteen vuoksi. Tutkimuksia kaupunkien elinympäristöistä ei ole vielä saatavilla.
Heimon historia
Nykyaikaiset muodot
| Liittyvien taksonien edustajat |
|---|


.jpg)
Tardigraden lähimmät sukulaiset löytyvät kahdesta hyvin erilaisesta ryhmästä: Niveljalkaiset , joihin kuuluvat äyriäiset (Crustacea), hämähäkit (Arachnida), tuhatjalkaiset (Myriapoda) ja hyönteiset (Insecta), ovat laajin eläinfyla, kun taas columbus (Onychophora), jota voidaan kuvata jaloiksi, muodostavat melko hämärän taksonin. Karhukaiset, niveljalkaiset, ja stumpworts hyvin todennäköisesti muodostavat luonnollisen perhe ryhmä yhdessä, ns monophyletic taksonin , joka tunnetaan Panarthropoda. Kehon segmentoitumista, joka esiintyy kaikissa tämän ryhmän eläimissä, ja pariksi muodostuneiden ruumiinlisäkkeiden ulkonäköä voidaan pitää yhteisenä johdettuina ominaisuutena; kutikularakenne on myös kaikkien kolmen taksonin yhteinen.
Panarthropodassa tardigrades on perinteisesti ryhmitelty kantojalkojen kanssa protoarthropodiksi:
| Panarthropoda |
|
||||||||||||
Alun perin ajatuksella oli rooli, että kolumbus ja tardigrade eivät ole vielä saavuttaneet niveljalkaisten organisatorista tasoa. Evoluutiotekniikassa kuitenkin katsotaan, että ero enemmän tai vähemmän pitkälle kehittyneiden muotojen välillä on vanhentunut; nykyaikaisten luokitusten tarkoituksena on vain heijastaa taksonien todellisia fylogeneettisiä suhteita toisiinsa.
Protoarthropodan yhteisiä piirteitä ovat pussimaiset kehon lisäosat, kantojen jalat, joita löytyy sekä kolumbuksesta että tardigradista. On myös useita muita homologioita , eli ominaisuuksia, jotka voidaan jäljittää yhteiseen esiasterakenteeseen. Erityisesti tardigrade -pää, joka oletettavasti koostuu kolmesta segmentistä, vastaa hyvin todennäköisesti kolobusirungon kolmea ensimmäistä yksikköä. Niiden korkokenkiä voidaan näin ollen pitää voimakkaasti muunnetuina toisen segmentin kehon lisäyksinä, ja ne olisivat siten homologisia samettimadojen leukoille. Kynsirakenne on myös hyvin samanlainen tardigrade ja columbus. Näistä silmiinpistävistä yhtäläisyyksistä huolimatta on kiistanalaista, muodostavatko Protoarthropodat luonnollisen perheryhmän, koska luultavasti kaikki luetellut piirteet ovat symplesiomorfioita , toisin sanoen ne löytyivät jo kaikkien Panarthropodan esi -isistä. 21. vuosisadan alussa mieluummin sisarryhmäsuhde tardigraden ja niveljalkaisten välillä syntyi:
| Panarthropoda |
|
||||||||||||
Tardigraden jatkosuhde nähdään perinteisesti annelidimatoissa (Annelida). Heidän kanssaan he jakavat muun muassa pehmeitä, nivelöimättömiä rungon lisäosia, joissa on lopulliset karkaistut "kynnet" ja päätyosa, joka sulkee vartalon eteenpäin. Tällä Articulata -hypoteesilla, joka juontaa juurensa ranskalaiseen luonnontieteilijään Georges Cuvieriin , oli edelleen paljon kannattajia 21. vuosisadan alussa , mutta se kyseenalaistettiin yhä enemmän 1990 -luvun loppupuolella morfologisten ja molekyyliperäisten löydösten vuoksi. Se korvattiin käsite, joka ryhmässä wormlike eläimiä, Cycloneuralia tarjoaa lähin sukulainen Panarthropoda: Tällä taksonin laskenta säiettä (Nematoda) ja string madot (jouhimadot), mutta myös kolme pikemminkin hämärtää eläinryhmät makkaramadot (makkaramadot), Koukku (Kinorhyncha) ja korsettieläimet (Loricifera). Panarthropodan tavoin kaikille näille taksoneille on tunnusomaista se, että ne hylkäävät niiden ei-solullisen ulkokuoren tai kynsinauhan ainakin niiden elinkaaren yksittäisissä vaiheissa; siksi niitä kutsutaan sulaviksi eläimiksi (Ecdysozoa):
| Sulavat eläimet (Ecdysozoa) |
|
||||||
Kuolleet muodot
Tardigrade -fossiilit ovat erittäin harvinaisia, joten ne eivät juurikaan auta ymmärtämään tämän eläinryhmän kehitystä. Joidenkin varhaisen geologisen aikakauden löydösten lisäksi tiedetään , että vain muutamia yksilöitä myöhäiseltä geologiselta ajalta on säilynyt keltaisena .
Lobopodit , ryhmä matoja, kuten Cambrian ja Ordovician geologisista aikakausista , jotka ovat peräisin edellisestä Ediacarianista , nähdään yhä enemmän Tactopodan, niveljalkaisista ja tardigradista koostuvan taksonin, mahdollisina edustajina . He juoksivat nivelöimättömillä, pussimaisilla tynkäjaloilla, jotka ovat samanlaisia kuin kantojalka, ja siksi ne on perinteisesti osoitettu tähän eläinheimoon. Jotkut nykyaikaiset kladistiset tutkimukset pitävät tätä arviointia perusteettomana ja pitävät sen sijaan lobododeja epäluonnollisena suhderyhmänä, josta sekä samettimadot että hyökkääjät ovat syntyneet. Ehdotettu vaihtoehto, joka ilmaisee tämän vaihtoehtoisen näkymän, on esitetty seuraavassa kaaviossa:
| Panarthropoda |
|
|||||||||||||||||||||||||||||||||||||||
Suvun Aysheaia , joka on vielä monia alkuperäisiä piirteitä kuten yksinkertainen rivi spesialisoituneen pitkät jalat tai sakkoa pinnallinen soittoäänen, joka ei vastaa huomaamaton kehon segmentointi, mikä muodostaa evoluution sisar ryhmä kaikkien muiden Panarthropoda, joka sitten jakaantuu stumpworts ja kaikki muut ryhmät hajoavat. Useita lobopods kuten raskaasti panssaroituja sukujen Cardiodictyon , hallucigenia tai Paucipodia oletettavasti muodostavat sammunut sivuhaaran, joka joutuu kuulumattomat suvun Luolishania toisaalta ja nimettömän taksonin lajista Kerygmachela kierkegaardi ja Tactopoda toisaalta.
Kerygmachela kierkegaardi , tämän hypoteesin mukaan Tactopodan välitön sisarryhmä, tunnetaan Pohjois -Grönlannin varhaisesta kambriasta ja on tardigradin ja niveljalkaisten tavoin selvästi myös segmentoitu ulkopuolelta. On epäselvää, kuuluvatko salaperäiset Anomalocaris -fossiilit myös Tactopodan varsilinjaan tai, kuten yllä on esitetty, liittyvät läheisemmin niveljalkaisiin kuin tardigradiin.
Ensimmäiset fossiilit, jotka voidaan selvästi osoittaa tardigradille, ovat peräisin Siperian Kuonamkan muodostumasta. Keski -kambrian 530 miljoonaa vuotta vanhassa kalkkikivessä on säilytetty neljä yksilöä, jotka voidaan tunnistaa kehon muodon ja koon perusteella tardigradiksi. Vielä kuvaamattomalla lajilla oli ilmeisesti kolme, ehkä jopa neljä jalkaparia, jotka päättyivät pariin epätasaisia kynsiä ja joiden kynsinauharakenne oli hyvin samanlainen kuin nykyisten eläinten. Eläimet , jotka ovat jo hyvin erikoistuneita, voidaan jopa nimetä johonkin nykyaikaisista luokista, heterotardigradasta .
Ei tiedetä, miten siirtyminen vankasta lobopodista alle millimetrin kokoiseen tardigradiin tapahtui. Yksi mahdollinen mekanismi olisi syntyminen , prosessi, jossa sukurauhaset muodostuvat ennenaikaisesti alkion kehityksen aikana ja siksi sukupuolinen kypsyys tapahtuu aikaisemmin kuin alkutila. Tämä pysäyttää nyt kehon kehittymisen ja erilaistumisen siten, että aikuisten eläinten kohdalla esiintyy toukkien tai nuorten ominaisuuksia, kuten merkittävästi pieni koko, ilmiö, jota kutsutaan pedomorfoosiksi .
Jopa silloin, kun tardigrade valloitti ensin maanpäälliset elinympäristöt, sitä ei voida sanoa varmasti fossiilisten tietueiden puutteen vuoksi. Koska tardigradea esiintyy nykyään usein sammaltyynyissä ja sammalit olivat hyvin todennäköisesti ensimmäisiä kasveja, jotka asuttivat maan ja joutuivat kuivumisriskin suhteen samanlaisiin haasteisiin kuin tardigrade itse, on hyvin mahdollista, että sammal ja tardigrades ottivat askel maihin yhdessä tarvittavien säätöjen kanssa.
Viimeistään tässä vaiheessa eläinten on oltava kehittäneet ainutlaatuisen sopeutumisensa kuivuuteen, joka tunnetaan kryptobioosina. Sen ansiosta he voivat yksinkertaisesti välttää haitallisia ympäristöolosuhteita sammuttamalla aineenvaihduntansa väliaikaisesti. Tästä seuraa, että ainakin ympäristön valinta on vain suhteellisen heikkoa, ja selittää siten fyysisen ulkonäön, fenotyypin, hyvin hitaan Bradytelin kehityksen .
Siksi ei ole yllättävää, että liitukaudelta saadut myöhemmät tardigrade -fossiilit eivät juurikaan osoita kehonmuutoksia nykypäivän lajeihin verrattuna. Beorn leggi, esimerkiksi laji on saatu Kanadan keltainen , voidaan jo osoitettu Eutardigrada luokassa; huonosti säilynyt nimetön nuori samalta paikkakunnalta voi kuulua luokkaan Heterotardigrada. Muita meripihkan fossiileja tunnetaan Yhdysvaltain New Jerseyn osavaltiosta , jotka oletettavasti ovat peräisin myöhäisestä liitukaudesta, nimeltään Turonian . Myös heidät on oletettavasti osoitettu Eutardigradalle, eivätkä ne salli muita johtopäätöksiä kuin että tämä kehityslinja on ollut olemassa yli 65 miljoonaa vuotta.
Ainoat muut tardigrade-fossiilit ovat hieman yli 7 000 vuotta vanhat munat, jotka ovat säilyneet Etelämantereen eteläpuolisissa turpeissa . Koska eri tardigrade -lajeilla on erilaiset lämpötila- ja kosteusasetukset, on harkittava eri suokerroksissa esiintyvien munien käyttöä, kuten kasvien siitepölyä, tuon ajan ilmaston määrittämiseen.
Järjestelmällisyys
Ei ole vakavaa epäilystä siitä, että tardigrades muodostaa monofyyttisen taksonin , eli palaa yhteiseen vanhempaan lajiin ja sisältää kaikki tämän lajin jälkeläiset. Tärkeitä synapomorfioita , yhteisiä, johdettuja piirteitä, ovat esimerkiksi teleskooppisesti sisäänvedettävät jalat ja suukappale.
Vuoteen 2005 mennessä on kuvattu noin 930 lajia , mukaan lukien 160 meren muotoa. Todellinen lajien määrä on luonnostaan tuntematon, mutta sen arvioidaan olevan noin 10 000. Lisäksi monet "lajit", jotka on rajattu morfologisten kriteerien mukaan, ovat oletettavasti piilossa salaperäisten lajien ryhmiä, jotka voidaan erottaa vain molekyyligenetiikan avulla, mikä lisäisi taksonin biologista monimuotoisuutta entisestään.
On kolme eri luokkaa, joiden väliset suhteet ovat edelleen epäselviä:
- Kuten heterotardigradaa kutsutaan "panssaroiduiksi" tardigradesiksi, vaikka kaikilla lajeilla ei todellakaan ole selkää ja ne ovat kovettuneet yksittäisiksi panssarilevyiksi (skleriiteiksi) jaettuina kynsinauhoina. Monissa muodoissa voidaan havaita silmiinpistäviä lisäyksiä, kuten cirri ja clavae; jalat voivat päättyä sekä kynsiin että liimalevyihin. Sukupuolielinten aukko on aina suoraan kehon pinnalla, Malpighian rauhasia erittymiseen ja osmoregulaatioon ei tapahdu. Heterotardigradaa esiintyy sekä meri- että raja -avaruudessa.
- Kuten eutardigrade kutsuu "alasti" vesikarhuja , niiden iho on ohut eikä kovettu. Huomattavia aistikarvoja pään päällä ei koskaan löydy tästä ryhmästä; jalat päättyvät pääosin kynsiin. Toisin kuin heterotardigrada, munanjohtimen putki avautuu peräsuoleen, josta tulee siten kloaka; eritystä Malpighian rauhasia käytetään erittymiseen. Suurin osa Eutardigradasta elää makeassa vedessä tai maalla, vaikka joitakin merilajeja on myös olemassa.
- Mesotardigrada tunnetaan vain yhdestä kadonneesta lajista, Thermozodium esakii , joka on virallisesti sijoitettu Thermozodiidae -perheeseen. Kuvauksen mukaan sen päässä on sivusuunnassa oleva sirkus, mutta ei clavae; suun ympärillä on neljä syylän kaltaista ulkonemaa (papillaa). Jalan pohjassa on myös papilloja, kun taas jalan pää sulautuu 6-10 yksinkertaiseen kynsiin; Malpighian rauhaset ovat läsnä. Laji löydettiin leväpehmusteista kuumalla lähteellä, tyyppisellä paikkakunnalla, lähellä Japanin Nagasakin kaupunkia , mutta se tuhoutui maanjäristyksessä toisen maailmansodan jälkeen. Koska mitään tyyppimateriaalia ei ole säilynyt ja lajia ei ole löydetty vasta tänään, sen olemassaoloa pidetään nyt kyseenalaisena ja sen nimi on siis substantiivi dubium .
Usein oletetaan, että yksi järjestys Heterotardigrada, valtaosaltaan meren Arthrotardigrada sisältää alkeellisin karhukaiset lajeja, joista muodoista muut Heterotardigrada järjestys Echiniscoidea ja erityisesti limnotardigrada perheen Echiniscidae on kehitetty, ennen kuin nämä vuorostaan muiden karhukaiset luokka Eutardigrada, jonka lajeja esiintyy pääasiassa makeassa vedessä ja maalla:
| Tardigrade |
|
||||||||||||||||||||||||
Tätä hypoteesia ei kuitenkaan ole vielä vahvistettu; alustavat molekyyligeneettiset tiedot viittaavat siihen, että sekä hetero- että eutardigrada liittyvät luonnollisesti toisiinsa:
| Tardigrade |
|
||||||||||||
Tutkimushistoria
Tardigradologia eli tardigrade -tutkimus on peräisin 1700 -luvulta . Saksalainen luonnontieteilijä Johann Conrad Eichhorn oli luultavasti ensimmäinen henkilö, joka havaitsi eläimiä 10. kesäkuuta 1767. Quedlinburgin pastori Johann August Ephraim julkaisi löydöksensä teoksessa Osallistuminen pienimpien vesieläinten luonnonhistoriaan, joita ei voi nähdä paljaalla silmällä ja jotka ovat Gdanskin ja sen ympäristön vesissä vasta vuonna 1775 ja näin vuosia myöhemmin . Goeze on enimmäkseen voimassa nykyään ryhmän löytäjänä. Hän voisi väittää hänen ensimmäinen havainto 10. joulukuuta 1772 julkaisi kuvauksen eläinten itsenäisenä kirjoitettuja muistiinpanoja sekä kääntää häntä Ranskan ja julkaistu 1773-lehdessä herra Karl Bonnets tutkielmia päässä Insektologie . Siellä hän kirjoitti mm.
”Tämä pieni eläin on outo, koska koko sen kehon rakenne on poikkeuksellinen ja outo, ja koska ulkoisessa muodossaan se ensisilmäyksellä muistuttaa eniten pientä karhua. Tämä myös motivoi minua antamaan hänelle pienen vesikarhun nimen. […] Ei kuitenkaan pidä pelätä miettiä näitä näkymättömän maailman saalistajia. "
Vuotta myöhemmin paluu anhydrobioottisesta tilasta havaittiin ensimmäistä kertaa, jota italialainen luonnontieteilijä Lazzaro Spallanzani kuvaili pian sen jälkeen "ylösnousemukseksi kuolleista". Juuri hän antoi ryhmälle nimen Il Tardigrada hänen kirjansa Opuscoli di Fisica animale e vegetabile , joka julkaistiin Modenassa vuonna 1776 , toisessa osassa . Vuonna 1790 eläimet sisällytettiin ruotsalaisen luonnontieteilijän ja systemaattorin Carl von Linnén teokseen Systema Naturae .
Ensimmäinen tieteellinen monografia ilmestyi vuonna 1840, yhdeksän vuotta myöhemmin löydettiin ensimmäinen meren tardigrade; Ensimmäisen fossiililajin, Beorn leggin , tieteellistä kuvausta joutui kuitenkin odottamaan yli vuosisata vuoteen 1964.
Eläinten järjestelmällinen sijainti oli epäselvä alusta alkaen; aikana koko 19th century, ne olivat taksonomisesti valinnaisesti rataseläimet (Rataseläimiä) tai meri robotit ryhmitetty (Pycnogonida) tai niveljalkaisten tarjotaan (Arthropoda). Jopa 1900 -luvun ensimmäisellä puoliskolla heidän tarkat filogeeniset suhteensa olivat kiistanalaisia, joten lopulta italialainen tardigradologi Giuseppe Ramazzotti korotti ryhmän oman eläinsuojelunsa arvoon. Taksonin lisäluokitus ulottuu saksalaiseen biologiin Ernst Marcusiin, joka jakoi vuonna 1929 kaikki tardigradit hetero- ja eutardigrada-alaryhmiin; Vuonna 1937 Gilbert Rahm esitteli Mesotardigradan kolmanneksi ryhmäksi.
Sekalaisia
- Israelin kuunlaskurin Beresheetin epäonnistunut laskeutuminen hajotti useita tuhansia tardigradeja. Koska he voivat pärjätä ilman happea ja heidät voidaan herättää henkiin lämpenemisen jälkeen jäädytetystä tilasta, heidät valittiin tähän kuutehtävään. ”Kuolleen materiaalin tavoin ne voivat selviytyä vuosia kuivuneena. Heidän olisi myös pitänyt selviytyä kuunonnettomuudesta ”, Nova Spivack vakuutti yhdysvaltalaiselle Wired -lehdelle . Nova Spivack on Arch Mission -säätiön johtaja.
kirjallisuus
- EE Ruppert, RS Fox, RD Barnes: Selkärangattomat eläintiede. Toimiva evoluutiomenetelmä. 7. painos. Brooks / Cole, Lontoo 2004, ISBN 0-03-025982-7 , s.510 .
- RC Brusca, GJ Brusca: Selkärangattomat. 2. painos. Sinauer Associates, Sunderland Mass 2002, ISBN 0-87893-097-3 , s.469 .
- IM Kinchin: Tardigradien biologia. Portland Press, Lontoo 1994, ISBN 1-85578-043-7 , s.186 .
- M. Blaxter, B. Elsworth, J. Daub: Laiminlyötyjen eläinsuojien DNA -taksonomia: tardigradien odottamaton monimuotoisuus. Julkaisussa: Proceedings of the Royal Society of London. Sarja B.London 271.2004, s. 189. ISSN 0375-0434
- GE Budd: "Opabinia regalis" -morfologia ja niveljalkaisten varsiryhmän rekonstruktio. Julkaisussa: Lethaia. Taylor & Francis, Oslo 29.1996, ISSN 0024-1164 , s.1 .
- RA Dewel, WC Dewel: Tardigradien paikka niveljalkaisten kehityksessä. Julkaisussa: RA Fortey, RH Thomas (Toim.): Niveljalkaisten suhteet. (= The Systematics Association Spec. Volume 55). Chapmann ja Hall, Lontoo 1998, ISBN 0-412-75420-7 , s.109.
- RA Dewel, DR Nelson, WC Dewel: "Echiniscus veridissimus" Peterfin aivot, 1956 (Heterotardigrada). Avain niveljalkaisten pään filogeenisen aseman ja kehityksen ymmärtämiseen. Julkaisussa: Zoological Journal of the Linnean Society. Blackwell, Oxford 116.1996, ISSN 1096-3642 , s.35 .
- JR Garey, DR Nelson, LY Mackey, L.Li: Tardigrade phylogeny, Congruency of morfologiset ja molekyylitodisteet. Julkaisussa: Zoologischer Anzeiger. Elsevier, Jena 238.1999, ISSN 0044-5231 , s.205 .
- A. Hejnol, R. Schnabel: eutardigrade Thulinia stephaniae on määrittelemättömän kehitys ja mahdollisuudet säädellä aikaisin blastomere ablaatioita. Julkaisussa: Development. 132.2005, s. 1349.
- A. Jörgensen, R. Kristensen: Tardigradan molekyylifylogeneesi - Heterotardigradan monofiilian tutkimus. Julkaisussa: Molecular Phylogenetics and evolution. Elsevier, Amsterdam 32.2004, 2, ISSN 1055-7903 , s.666 .
- DR Nelson: Tardigrada. Julkaisussa: JH Thorp, AP Covich (Toim.): Pohjois -Amerikan makeanveden selkärangattomien ekologia ja luokittelu. Academic Press, San Diego Ca 2001, ISBN 0-12-690647-5 , s.527 .
- DR Nelson: Tardigraden nykyinen tila. Evoluutio ja ekologia. Julkaisussa: Integrative and Comparative Biology. Lawrence 42,2002, ISSN 1540-7063 , s. 652.
- DR Nelson, NJ Marley: Lotic Tardigradan biologia ja ekologia. Julkaisussa: Makean veden biologia. Blackwell, Oxford 44.2000, ISSN 0046-5070 , s.93 .
- Joh. August Ephraim Goeze: Herra Karl Bonnetin tutkielmat hyönteistieteestä. (PDF; 32,7 Mt). Täydennys kääntäjän JAE Goeze, Halle 1773. (Alkuperäinen teksti: Tämä pieni eläin on outoa, koska koko rakenne sen ruumis on poikkeuksellista ja outoa, ja koska sen ulkoisen muodon, ensi silmäyksellä, näyttää kaikkein kuten pieni karhu on. Ss 367–375)
nettilinkit
- Tardigrada Tähtitiede Päivän kuva on NASA (6. maaliskuuta 2013 Englanti )
- Tardigradan uutiskirje (eng.)
- Tardigrades: valokuvat ja videot (englanti)
- Laajat tosiasiatekstit, valokuvagalleriat ja lyhytelokuvat tardigradesta
- Taksonomia suvun tasolle asti
- Mikroskopia Iso -Britannia
- Tutkimus tardigrade avaruudessa
- Sveitsiläinen tardigradien ekologian, fysiologian ja evoluutiobiologian tutkimuskeskus
- Äskettäinen väite, jonka mukaan tardigrades sai kuudesosan DNA: staan mikrobeista, on alkanut purkaa (Engl.)
- Claudia Ruby: Tardigrade - Pehmeä selviytymistaiteilija Baijeri 2 Radiotieto . Lähetys 25. heinäkuuta 2019 (podcast)
- Mutta ei tuhoutumaton - se on tardigraden heikko kohta
- Lars Fischer: Sininen hehku suojaa tardigradia säteilyltä , osoitteessa Spektrum.de 15. lokakuuta 2020
- Peter Dockrill: Olemme juuri löytänyt toinen temppu karhukaiset Käytä pohjimmiltaan Indestructible puolesta: tiede hälytykseen 14 lokakuu 2020
Yksilöllisiä todisteita
- ↑ Andreas Hejnol, Ralf Schnabel: Eutardigrade Thulinia stephaniaella on epämääräinen kehitys ja potentiaali säätää varhaisia blastomere -ablaatioita. Julkaisussa: Development. 132, 2005, s. 1349-1361. doi: 10.1242 / dev.01701
- ↑ Daiki Horikawa et ai.: Säteilytoleranssi tardigrade Milagnes tardigradumissa . Int. J. Radiat. Biol. 82, 2006, doi: 10.1080 / 09553000600972956 ( ilmainen koko teksti ).
- ↑ Emma Brennand: Tardigrades: Vesi karhuja avaruudessa . Toimittaja: BBC. 17. toukokuuta 2011 ( bbc.co.uk [käytetty 30. marraskuuta 2019]).
- ↑ Peter Reuell: Harvardin tutkimuksen mukaan asteroideilla voi olla avainrooli elämän levittämisessä. Julkaisussa: Harvard Gazette. 8. heinäkuuta 2019, Haettu 30. marraskuuta 2019 (amerikkalainen englanti).
- ↑ Lorena Rebecchi et ai. : Anhydrobioottisen eutardigrade Paramacrobiotus richtersin vastustuskyky avaruuslennolle (LIFE-TARSE-tehtävä FOTON-M3: lla) , julkaisussa: Journal of Zoological Systematics and Evolutionary Research 49, 21. huhtikuuta 2011, s. 98-103, doi: 10.1111 / j. 1439-0469.2010.00606.x .
- ↑ Neil A. Campbell et ai. : Biologia. 8., päivitetty painos. 2011, ISBN 978-3-8273-7287-1 .
- ↑ D. Schulze-Makuch et ai. : Tardigrades: Esimerkki monisoluisista ekstremofiileistä. Julkaisussa: Joseph Seckbach et ai.: Polyekstremofiilit - elämä monenlaisissa stressimuodoissa. Springer, Dordrecht 2013, ISBN 978-94-007-6487-3 , s. 597-607.
- ↑ Thomas C.Boothby, Hugo Tapia, Alexandra H.Brozena, Samantha Piszkiewicz, Austin E.Smith: Tardigrades käyttää luontaisesti häiriintynyttä proteiinia kuivumisen selvittämiseksi . Julkaisussa: Molecular Cell . nauha 65 , ei. 6. maaliskuuta 2017, ISSN 1097-2765 , s. 975-984.e5 , doi : 10,1016 / j.molcel.2017.02.018 , PMID 28306513 ( cell.com [näytetty päivänä kesäkuuta 20, 2017]).
- ↑ D. D. Horikawa, T. Kunieda et ai. : Perustetaan äärimmäisen siedettävän tardigradin Ramazzottius varieornatus -kasvatusjärjestelmä: uusi astrobiologian mallieläin. Julkaisussa: Astrobiology. Osa 8, numero 3, kesäkuu 2008, s.549-556, doi : 10.1089 / ast.2007.0139 , PMID 18554084 .
- ↑ Fumihisa Ono et ai. : Erittäin korkean paineen vaikutus pieneläimiin, tardigradesiin ja Artemiaan.Cogent Physics 3, 2016, doi: 10.1080 / 23311940.2016.1167575 (ilmainen koko teksti).
- ↑ B. Vargha, E. Otvos, Z. Tuba: Tutkimukset ekologisia vaikutuksia raskasmetallit Unkarissa sammal-asunnon veden kantaa (karhukaiset), kuten bioindikaattoreina. Julkaisussa: Ann Agric Environ Med. 2002; 9 (2), s.141-146. PMID 12498580
- ↑ Klaus J. Müller, Dieter Waloszek, Arcady Zakharov: 'Orsten' -tyyppinen fosfatoitu pehmopaperin säilyttäminen ja uusi ennätys Siperian Keski-Kambrian Kuonamkan muodostumasta. Julkaisussa: New Yearbook for Geology and Paleontology. Papers 197 (1), 1995, s. 101-118.
- ↑ J. A. E. Goeze: Herra Karl Bonnetsin tutkielmat hyönteistieteestä. J. J. Gebauer, Halle, 1773. Digitoitu versio katsottu 9. syyskuuta 2016. Lainaus osoitteesta www.baertierchen.de .
- ↑ Patrick Illinger: Tardigrades ovat kuun ensimmäiset asukkaat . Julkaisussa: Süddeutsche Zeitung , 7. elokuuta 2019